心 理 学 报 2004,36(2):219~234 ActaPsychologicaSinica
顿悟的大脑机制*
罗 劲
(中国科学院心理健康重点实验室,北京100101)
摘 要 自从柯勒1917年提出顿悟的概念以来,这个问题一直吸引着心理学家的关注。但有关顿悟过程的精确的大脑机制却始终未被触及。从心理过程上看,顿悟是一个瞬间实现的、问题解决视角的/新旧交替0过程;它包含两个方面,一是新的有效的问题解决思路如何实现,二是旧的无效的思路如何被抛弃(即打破思维定势)。我们以谜语作为材料,利用功能性磁共振成像(fMRI)技术精确记录了人类的大脑在实现顿悟的一瞬间的活动状况。结果显示顿悟过程激活了包括额叶、颞叶、扣带前回以及海马在内的广泛脑区。根据各方面的综合证据,该文认为:顿悟过程中,新异而有效的联系的形成依赖于海马,问题表征方式的有效转换依赖于一个/非语言的0视觉空间信息加工网络,而思维定势的打破与转移则依赖于扣带前回与左腹侧额叶。关键词 顿悟,事件相关fMRI,问题解决。分类号 B842
1 引言:顿悟研究的一般背景
顿悟(insight)的研究始于格式塔心理学家柯勒,它挑战了当时占主导地位的桑代克的/尝试-错误0学习理论,证明问题解决过程可以以/突变0而不是/渐变0的方式发生,因而具有重要的理论意义。但柯勒关于顿悟的研究的意义却并不止于对于/尝试-错误0理论的反驳,它的意义还在于采用科学心理学的方法界定了一个高级的认知过程)))顿悟,顿悟过程是我们的创造性思维赖以实现的基础。 顿悟式的问题解决含有/突发性0(suddenness)、/直指性0(directness)和/(行为操作的)持续性0(con-tinuousnessofperformance)三个特征[1],/突发性0是指顿悟式的问题解决往往在极短的一瞬间突然实现;/直指性0是指有效解决问题的方案在顿悟状态中直接呈现在眼前,并非经过反复的尝试错误或者复杂的手段-目的分析;/持续性0是指顿悟一旦获得,便很少出现行为上的反复,它很像一种/一学即会0的学习过程。经典的顿悟问题包括/六火柴问题0,/蜡烛问题0,/双绳问题0,和/九点问题0等。一般而言,顿悟问题包含三个显著的特征,其一,顿悟问题的解决并不需要特殊的认知技能;其二,人们在解决顿悟
问题的过程中,往往会遇到一个明显的障碍,这个障碍让他们感到不知所措;其三,这个障碍一旦被打破,顿悟问题就会迎刃而解,人们会有一种伴随情感释放的/啊哈0的体验[2]。
自从柯勒1917年提出顿悟的概念以来,这个问题一直吸引着心理学家的关注。但是20世纪50年代认知科学兴起之后,有关顿悟的研究有近十数年的沉寂,其原因可能与以口语报告法以及/手段-目的0分析为代表的问题解决实验研究范式的兴起有关。手段-目的分析注重问题是如何在头脑中被解决的,而非人们的头脑如何表征问题,因此,手段-目的分析更为关注一个逐渐趋近目标的/渐变0过程而非/突变0的顿悟过程。到了90年代,有关顿悟的研究开始回潮[3],这可能与80年代以后,认知科学研究中的主流范式逐渐淡化,研究的领域趋于多元,以及当前社会对创造性思维的日益关注有关。 目前,有关顿悟的心理学理论较多,但从总体上可分为两大类。
一类理论着重于探讨顿悟中新的、能有效地解决问题的思路如何实现,比如/格式塔的知觉场理论0认为顿悟源于知觉场的重新组织,在问题尚未解决之时,知觉场处于一种紧张的不平衡的状态,而一
收稿日期:2003-08-11
*
本研究受国家自然科学基金(批准号30270464)、FAIPICS以及留学回国人员科研启动基金(教外司留[2002]247号)资助。
通讯作者:E-mail:luoj@psych.ac.cn
220心 理 学 报36卷
旦一种均衡良好的知觉场被形成,问题就迎刃而解了,而良好均衡的知觉场的形成取决于问题解决者超越了对于事物的表面特征的认识,而领悟到事物之间的内在关系[4,5]。/表征变换理论0认为通常顿悟问题会引导人们形成不适当的问题表征从而阻碍人们有效地解决问题,成功的问题解决取决于问题表征方式的变换[6~
9<
并逐步修改这些思路以使之适应当前的问题情境0[14]。值得一提的是,史学研究资料表明,巴甫洛夫在1933~1936年期间,也作了一系列以猩猩为被试的实验,重复了苛勒关于顿悟的发现,但是,巴甫洛夫倾向于用条件反射的概念解释顿悟现象,他也认为顿悟是逐渐获得的而不是突然获得的[15]。 尽管目前已经建立了多种不同的有关顿悟的心理学理论,但迄今为止,有关顿悟的大脑机制问题却始终未被触及。作为一个突发的、直指人心的问题解决过程,顿悟无疑是由一个强大的脑动力过程来实现的,这个过程是怎样的?由哪些关键的大脑神经结构参与了这个过程的实现?这些神经结构与怎样的认知心理过程相对应?对上述问题的回答,无疑将对我们认识顿悟的本质,揭示创造性思维的脑神经机制开启良好的开端。
[16]
;但问题的表征方式并不是
轻易就可以被变换的,这常常需要一个组块的破解过程(chunkdecomposition)。组块是人们在日常生活过程中逐渐形成的、由各个不同的要素所构成的紧密的统一体,组块内诸要素的紧密程度将决定问题表征方式是否能够被有效地变换。/类比理论0认为顿悟问题的解决关键在于,问题解决者通过一个类比过程,以创造性的方式将原本不直接相关的思路或者方法应用于当前的问题情境[5],比如,用骑士进攻城堡的情景来类比放射治疗肿瘤的手段。
另一类理论着重于探讨旧的思维方式何以会阻碍问题的有效解决。比如/机能固着理论0认为人们对一个特定物体的表征,往往与这个物体的通常功用联系在一起,当一个熟悉的物体将被以一种不熟悉的方式使用时,就会发生困难[10]。/心理成规理论0认为人们会反复地在那些无效的思路上进行尝试,这种徒然的努力会起到相反的作用,它使得这条错误通路的激活程度越来越强,并同时降低了问题解决者探索其它有效途径的可能性[11],这个理论可以被形象地理解为强调一种问题解决过程中的/强迫症0倾向的假设。/心理定势理论0认为思维习惯会使问题解决者不再致力于寻找新的、可能是更为有效的问题解决途径[12],典型的例子是著名的Luchins的/水罐实验0。/心理动力学理论0认为顿悟问题往往有赖于一个/迂回0的解决方案。但是,由于动机的驱使,问题解决者通常会尝试直接的途径,而这种途径是无效的[13]。比如,该理论认为/九点问题0之所以困难,并不是因为问题解决者的思路局限于九个点所限定的框架,而是因为问题解决者总是试图用一条直线去连接尽可能多的点。 除了上述两个大类理论以外,还存在一些其它有关顿悟的心理学解释。但从严格的理论角度来考虑,这些解释是反顿悟的,因为它们认为顿悟并无特殊之处(nothing-new),顿悟与普通的学习过程并不存在实质性的差别。基于过去的经验与顿悟之间存在密切的关系,Weisberg指出:/几乎没有什么理由相信顿悟是一种与过去经验毫无关系的灵光一现0,/,
2 顿悟的脑成像研究
新近发展起来的脑成像技术(如fMRI,PET,EEG等)为直接观察脑在处理复杂信息时的活动状况提供了强有力的研究手段。形象地说,这些技术使得大脑的/黑箱0变得/透明0。特别是事件相关功能性磁共振成像技术(event-relatedfMRI)使得我们能够以毫米水平的精确程度记录大脑在一个瞬间的活动状况,从而为研究顿悟的大脑机制提供了适合的技术手段。尽管如此,对于顿悟现象的研究仍然有一定的难度。因为顿悟有别于通常的思维过程譬如加减乘除运算,顿悟是一个较为罕有而又瞬间即逝的/发现0状态。采用脑成像技术研究顿悟的脑机制主要有两个困难,其一,在自然状态之下,顿悟发生的时间点是不可控的,有研究表明:人们对于顿悟问题的思考和解决需要短到数秒,长到数天甚至数月的时间,而实验室条件下的脑成像扫描只能有10~20分钟的时间,如何在如此短暂的时间内获得顿悟过程?其二,从脑成像技术上考虑,可靠的分析需要通过对相当数量的、可重复观察的同类心理事件进行叠加才能够获得,而经典的实验室条件下所使用的顿悟问题(如六火柴问题,蜡烛问题,双绳问题,九点问题等)只有为数不多的几个,并且这些问题的性质、特征、复杂程度等也都各不相同,因而难以作为脑成像研究的实验材料。针对上述困难,我们在最近的研究中采取了两项措施,一是采用传统的谜语作为实验材料,二是通过向被试呈现标准答案来/催化0顿悟过程。谜语材料的资源远比经典的顿悟[16]
2期罗劲:顿悟的大脑机制 221
的谜语作为实验材料(谜语的例子:/你杀死了她,但却得流你自己的血0)))谜底:蚊子)。通过一个预备实验的筛选,我们选择了那些被试能够良好地理解、但仍不知其答案的谜语作为实验材料(因此,每个被试的实验材料都适合于这个被试本人的情况)。在扫描过程中,先向被试呈现那些事先选择好了的谜语,令其进入对特定问题的准备和思索状态,然后突然呈现谜语的答案,被试在看到答案的瞬间便会产生顿悟。这样,我们就在一个有限的扫描时间之内获得足够数量的顿悟的心理事件,并且可以确切地锁定顿悟的心理事件发生的时间,从而确定那个时刻点上发生了怎样的大脑活动。具体的实验是利用一台磁场强度为3.0特斯拉的核磁共振成像设备完成的,我们记录了七名大学生被试在顿悟性地解决谜语时的整个大脑的活动状况。分析结果表明:相对于静息状态,人脑在顿悟过程中有广泛的脑区被激活,包括双侧的额上回、额中回和额下回,扣带前回,双侧的颞上回及颞下回,以及楔前叶和海马等脑区。从我们目前所掌握的文献来看,这项研究是历史上首次精确地记录人类大脑在顿悟时的活动状况的研究报告。我们基于以下的三个起点对于这个结果进行讨论:第一,顿悟过程不大可能由一个脑区完成,它是多个脑区共同完成的;第二,脑成像研究在本质上是一种相关的研究,我们通过脑成像研究所确定的是与顿悟过程相关联的脑区,但脑成像研究原则上并不能提供心与脑在因果关系上的推论;第三,参与顿悟的诸多脑区的作用不大可能是等同的,一些区域的作用会比另一些更加关键,要想对这些区域的贡献加以区分,就需要来自/领域内的证据0)))操纵不同的实验条件来分析某些功能和/跨领域的证据0(cross-domainvalidity))))即来自其他相关领域的佐证。
从心理过程上看,顿悟是一个在一瞬间实现的、问题解决视角的/新旧交替0过程,它包含两个方面,一是新的能有效地解决问题的思路如何实现,二是旧的无效的问题解决思路如何被抛弃(即思维定势如何被打破)。本文将着重讨论从这两个方面探讨顿悟的大脑机制。
[16]
的箱子叠加在一起才能取食到香蕉。在这项顿悟式的问题解决中包含了两个核心的技能,一是将箱子叠加在一起,二是爬到一个物体(比如箱子)上去够取另外一个物体(香蕉)。事实上,这两项技能都是黑猩猩所熟知的。因此,黑猩猩的顿悟行为,乃是有效地将这两项技能在一个特定的问题情境之下联结起来。直到1984年,这一设想才由Epstein等人在Nature杂志上发表的一项动物实验所证实[1]。实验者在充分消除了鸽子的飞行行为之后,训练鸽子获得了两项技能,一是去啄一个可以使之获得食丸的按钮,二是推动一只小木箱到特定的位置。研究者观察到:当按钮的位置被安置得比较高,鸽子够不到时,它就会将事先放在笼子另一边的小木箱推到按钮的下方,并爬上木箱去啄按钮,而鸽子的这种行为具有明显的顿悟的行为特征,即表现出/突发性0、/直指性0和/(行为操作的)持续性0。Epstein等人的实验说明了顿悟是旧有技能的创造性组合和联结。
当今认知神经科学普遍认为,在大脑中负责形成新异联系的关键性脑结构在海马[17~19]。有关海马的机能的一个典型的例子是著名的海马损伤病人H.M.,他在其他的认知机能(比如注意、工作记忆,语义记忆,思维和推理等)保持相对完好的情况下,唯独不能形成情节记忆。有趣的是,对H.M.个案的考察中发现:H.M.病人没有顿悟和/啊哈!0(aha)感。研究者使用的是一些含有歧义词的句子,比如/船长很喜欢自己的新位置0,在这句话中,/位置0(position)一词有两个意思,它既可以指一个具体的物体比如椅子,也可以指一个抽象概念比如船长这一职位。被试被要求确定这个句子的双重含义。正常被试在成功地发现了双重含义时,会表现出一种认知与情绪上的释放感,即表现出/啊哈0反应。而H.M.没有这种/啊哈!0反应。即使他偶尔成功地找到第二重的含义,他的反应也十分平淡,似乎并不认为这是一种成功的解决。这一早期的观察为海马在/啊哈!0反应以及顿悟中的作用提供了直接的脑损伤的依据。
与上述的推测相一致,我们在顿悟中观察到了明显的海马的活动(图1A)[16]。我们认为,实验中所使用的传统谜语的构成形式,通常是用一种不常见的表达方法描述一种常见的事物,比如将人拍死蚊子的行为描述为/你杀死了她,但却得流你自己的血0。而对于这个问题的顿悟性的解决会包含着一[20]
3 参与新异联系形成与问题表征转换
的脑机制
3.1 新异而有效的联系的形成与海马
顿悟意味着新异而有效的联系的形成。在柯勒,
222心 理 学 报36卷
不常见的描述方式联系在一起。我们认为,在顿悟中观察到的海马的活动,并不是由于词(标准答案)的新异性所引起的。以往的研究表明:海马会为新异的刺激所激活[21~
24]
所涉及的都是日常生活中极为常见的概念(如蚊子、河流等),这样的常见词不可能具有足以激活海马的
新异性。
。但是在这项研究中,谜语
图1 海马在顿悟中(A)与任务相关联系形成中(B)的活动(箭头所指的白色区域代表脑区的激活范围)
A为顿悟状态与静息状态相比较/减所获得的结果,被试数:7;差异显著水平:P<0.001(SPM{T},conjunctionanalysis).
B为对于任务相关词对的加工与对于任务无关词对的加工相比较/减的情况,被试数:7;
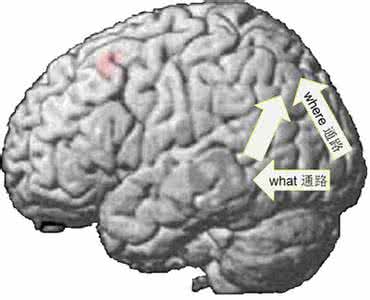
差异显著水平:p<0.001(SPM{T},conjunctionanalysis).
在Redish有关海马的功能的模型中包含了一个叫做参照框架(referenceframe)的子系统[25],这个子系统的活动依赖于动物当前的目标或者任务,换言之,参照框架体现了动机的作用。这个假设的主要证据来自于对海马内/目标-趋近0细胞的研究。1987年,Eichenbaum等人观察到海马内存在一种/目标-趋近0细胞(0goal-approach0cells)[26],这种细胞的活动特征与以前观察到的位置细胞(placecell)有所不同,位置细胞在动物一旦到达某个特定的位置就活动,而/目标-趋近0细胞与位置细胞的不同在于(1)/目标-趋近0细胞的活动与动物当时所处的位置关系不大;(2)/目标-趋近0细胞的活动具有前瞻性,它往往在到达特定目标以前2秒就开始活跃;(3)/目标-趋近0细胞在动物行动时就开始活动而不是在动物到达某个位置时活动;(4)只有当某个特定的位置是通向目标(食碗)的必经之路时,这个位置才能激活/目标-趋近0细胞,否则,即使动物经过这个位置,/目标-趋近0细胞也不活跃;(5)/目标-趋近0细胞只有在动物走向食碗并有食物可吃时才活动,它并不对食碗的位置本身起反应,也不在无强化的系列中活动。上述/目标-趋近0细胞的存在据此,我们假设:海马并不是在盲目地形成新异联系,相反,海马是在形成与当前的任务密切相关的/任务相关联系0(task-relatedassociation)[27]。这一假设的主要证据来自于对于Henke等人实验观察的反驳。Henke等人的实验旨在证明海马在形成新异联系中的作用[28]。他们给被试呈现一个无关的词对,比如/levelneed0,一种加工条件是要求被试分别判断自己是否喜欢这两个词的意义,而另一种加工条件则是要求被试判断这两个词的意义是否匹配,因此,上述两种条件都属于深度加工,而它们的不同在于后者是联系性的,它要求把两个无关词联系起来,而前者则不是联系性的,它只涉及两个词的独立加工。脑成像实验表明,只有在联系性加工的条件下,海马才活动。这个实验的意义在于澄清了一般意义上的深度加工与联系加工之间的区别。但是我们认为:Henke等人的实验中所采用的词义的喜好判断与词义的匹配判断可能并不等同,它们可能涉及不同的加工过程,因此,可能存在变量上的混淆。我们采用三词判断的方法,给被试呈现三个词,上面一个,底下两个,要求被试判断下面的两个词中哪一个在语义上与上面的词较为匹配。共有三种实验条,/00,
2期罗劲:顿悟的大脑机制 223
无明显的语义联系(如,上面的词为/土地0,下面的词为/钟表0和/音乐0);在/1关系0条件下,下面的两个词中有一个与上面的词有明显的语义联系,而另一个则没有(如,上面的词为/土地0,下面的词为/建筑0和/音乐0);在/2关系0条件下,下面的两个词都与上面的词有明显的语义联系(如,上面的词为/土地0,下面的词为/建筑0和/田野0)。我们假设:如果海马的作用在于形成新异的联系的话,那么,在三种条件下,海马的激活水平应该依次为/0关系0>/1关系0>/2关系0;但如果海马的作用在于激活旧有联系的话,那么,海马的活动平应该依次为/0关系0</1关系0</2关系0。研究的结果揭示了后一种情况,因而不支持Henke等人的结论[27]。那么,为什么海马在旧有联系的激活中活动更为强烈?这与海马参与/任务相关联系0的形成有关,在本实验中,信息加工的任务就是寻找两个词在语义上的相关性,因此,相关的词对越多,形成的/任务相关联系0就越多,海马的活动也就越加强烈。
为了进一步澄清/任务相关联系0与词对的语义相关性之间的关系,我们给被试呈现一个包含有各种不同的语义关系的词对的词单,在这个词单中,有的词对没有明显的语义关系,如/大象-茶叶0;有的则包含有一般意义上的语义关系,如/钢笔-纸张0;有的则包含有特定的语义关系,如范畴-例子关系(/家具-桌子0)和相对关系(/困难-容易0)。实验的关键设计在于,在一个区组中,我们要求被试判断每个词对是否包含范畴-例子关系,而在另一个区组中则要求他们判断每个词对是否包含相对关系,这样,相同性质的词对(比如包含相对关系的词对)在一种条件下是/任务相关0的,而在另一种条件下则是/任务无关0的。通过比较相同性质的词对项目在任务相关与无关的条件下对海马的激活情况,我们进一步证明了海马只有在加工与当前任务有关的关系时才活动(图1B),我们的观察也同时表明了词对的语义相关性本身并不足以激活海马*。如果将海马在任务相关联系的形成中的作用与海马在长时记忆中的作用联系起来加以考虑,就有可能从一个更为普遍的进化论的角度考虑海马的作用,任务相关联系对于生物个体的生存具有重大意义,海马对于这些联系的形成起反应并且将其保持在长时记忆之中,这无疑会增加生物个体的生存机会。上述讨论意味着,作为一个非常显著的任务相关联系
*
的形成过程,顿悟的发生需要海马的参与。 作为一个具有重要功能、而又结构相对简单、与其他大脑区域之间的联系相对明确的神经结构,海马结构在许多不同的物种中都可以被良好地界定,这也就是这个神经结构被广泛地研究的原因。目前,有关于海马的研究从总体而言有两大类,一类是动物实验,主要研究海马的损伤对于动物的找路行为的影响,或者记录海马中的神经元在动物的找路行为中的活动状况。另一类是人类实验,主要是研究海马与情节记忆或者陈述性记忆的关系。Redish认为,海马的这两类功能实际上是统一的,海马的作用在于不断地校正在自我导向系统中发生的错误,回忆情景以及填补情景上的空缺[25]。其中,最突出的例子是动物的重新定向行为(指动物在迷路以后的找路行为)。研究表明:在有明显的清晰可辨的路径标志的情况下,即使是海马被损伤的动物,也有可能从迷失的道路上找回;但是,当缺乏明显的外在的标志时,海马损伤的动物就难以找回原路。在后一种情况下,动物只有将自己的动作(比如跑了多远的路程)与某些外在的信息有效地综合起来,才能成功地实现自我导向。顿悟无疑包含着思维的重新定向过程,海马参与顿悟的事实提示:人类思维的重新定向可能与其空间定向的过程共享一个神经机制。与此设想相一致,新近脑成像的研究采用电脑虚拟空间研究了人类被试在执行托尔曼式的认知地图任务中大脑的活动,结果发现:相对于走老路时的脑活动状况,当人们去利用一条虽然已知但却从未走过新路趋近目标时会有海马的参与,其活动也在右侧,与我们在顿悟中观察到的海马的活动相一致。这一思考涉及人们在顿悟中的问题表征方式如何转换。
3.2 顿悟中问题表征方式的转换与脑内视觉空间
信息加工网络
问题表征方式是问题解决中的一个关键问题。Simon认为:/解决一个问题,仅仅意味着把这个问题表征到能够使答案变得明朗的程度0[30]。有关顿悟的表征变换理论认为:顿悟问题会引导人们形成不适当的问题表征从而阻碍人们有效地解决问题。成功的问题解决取决于问题表征方式的变换[7,8]。那么,这种变换是如何实现的?它是以一个线性言语的方式抑或是以一个空间视觉的方式实现的?以往有关顿悟的研究表明:顿悟可能是以一种非言语的
[29]
[16,27]
LuoJ,NikiK.Hippocampus.sroleinprocessingtask-relatedknowledge:flashingtothethingyouarelookingfor.Submittedmanuscripts.
224心 理 学 报
*
36卷
空间视觉的方式实现的。概括而言,其证据有以下几种[2]。第一,人类历史上的那些伟大的发明或者发现的提出过程往往是非言语性的,对此,詹姆斯总结说:/伟大的思想家会预兆性地在一瞬间窥见事物之间的全部关系,整个过程发生的速度是如此之快以至于无法言表0,与此相应,爱因斯坦也说:/那些思想不是以语言的形式来临的,我极少用语言来思考,一旦思想来临,我事后也许会想到要用语言去表达它0(转引自文献[2])。第二,实验室条件下的观察也显示:在顿悟来临之前,被试往往会有短暂的沉默;例如,Darkin要求被试在解决顿悟问题的时候口语报告其思维过程,结果发现:被试在发现关键性步骤时都没有言语活动[31]。第三,尽管被试的知道感判断(FOK)可以预测常规的推理问题的解决,但它却并不能预测顿悟问题的解决[32~
34]
一项研究中比较了两类问题的解答,一类是/脑筋急弯问题0,另一类是普通的百科知识性问题(比如/老话说:-男女七岁不能,,.不能怎样?0)))答案:不能同席)。在大学生被试事先对于这两类问题的答案都不知道的情况下,给他们看标准答案。结果表明:相对于顿悟的问题解决,百科知识问题的解决激活一个以左侧前部颞中回(布罗德曼21区)和左侧岛叶(布罗德曼13区)为中心的/知识语言信息加工网络0,而相对于百科知识问题的解决,顿悟的问题解决则激活一个包括双侧的后部颞中回(布罗德曼39区)、枕中回(布罗德曼19区)、楔前叶(布罗德曼19区)以及左侧海马旁回在内的/视觉空间信息加工网络0(图2A)。
这一结果与Goel等人有关三段论推理的研究是一致的[37]。在Goel等人的脑成像研究中,他们比较了两类三段论推理的脑过程,一类是包含具体名称的推理,比如,/所有的狗都是宠物,所有的卷毛狗都是狗,所有的卷毛狗都是宠物吗?0;另一类是只包含抽象符号的推理,比如/所有的P都是B,所有的C都是P,所有的C都是B吗?0。研究结果表明:与抽象的三段论推理相比较,具体的推理激活一个/知识语言信息加工网络0,而与具体的三段论推理相比较,抽象的推理激活一个/视觉空间信息加工网络0。Goel和Dolan认为,抽象的三段论推理之所以会激活一个/视觉空间信息加工网络0是因为在没有具体实物的情况下,人们在其头脑中可能会用圈图来表征包含、相交或者分离的关系,这样就会激活/视觉空间信息加工网络0[38]。
但这样的解释并不适用于我们的实验,因为在我们的实验中并没有涉及抽象关系,所有的名词都是具体的。一种可能性是在/脑筋急转弯问题0的解决中包含了某些特殊的形象,比如毛毛虫与蝴蝶,乃是这些可视的形象激活了视觉空间信息加工网络。但这样的假设与以下的两个事实不相符合:其一,这个实验所使用的百科知识性问题的可视觉化程度与/脑筋急转弯问题0相当,比如/男女七岁不同席0也是可以视觉化的。其二,我们进一步在两类/脑筋急转弯问题0之间进行了比较,一类的问题的视觉化程度较高,比如/夜里,一个穿白长袍的女人在海边的沙滩上行走,可她的身后却没有脚印,这是为什么?0(答案:她在倒退着走);另一类则视觉化程度较低,比如/诸葛亮是大智者,如果他还活在世上,我们现
,也就是说,被
试对于他们能否顺利解决所面临的问题事先一无所知,解决是突然发生的。第四,Maier以看似无意的方式向被试提供了解决顿悟问题的暗示,结果发现,那些能够抓住这个暗示、并利用它顿悟地解决问题的被试反而不能在意识里明确地感到暗示的出现并从中引申出其作用[35]。第五,Schooler,Ohlsson和Brook的研究表明:口语报告不但不能促成顿悟,反而会抑制顿悟过程。总体上讲,有关信息加工的认知理论可分为两大类,一类假设信息是以连续的方式被加工的,另一类则假设信息是以间断的(全或无的)方式被加工的,利用SDA(speed-accuracyde-composition)技术,Smith和Kounios证明:与一般的任务不同,顿悟问题是以全或无的方式解决的,亦即在顿悟中信息加工过程是不连续的,在没有阶段性的部分信息产生的情况下,顿悟会突然发生,这就决定了顿悟的非言语特性[36]。
为了研究顿悟中的问题表征方式如何转换,我们采用脑筋急转弯问题(日本称/脑的体操0)作为实验材料,脑筋急转弯问题与传统的谜语的一个最大的不同之处在于,传统谜语所描述的问题情境常常比较模糊(因此,在传统谜语中出谜者常常会为答案制定一个范围,如/打一体育用品0等),而脑筋急转弯问题则对问题情境有清晰的界定(比如,/有一条毛毛虫想过河,但河宽水深,又没有桥和渡船,也没有谁能帮助它,请问这条毛毛虫如何渡河?0)))答案:变成蝴蝶飞过河),因而,采用脑筋急转弯问题有利于探讨顿悟中的问题表征方式如何转换。我们在
*
[2]
LuoJ,NikiK,PhillipsS.Insight:the0internalsight0ofthesolution.Submittedmanuscripts.
2期罗劲:顿悟的大脑机制 225
在的世界一定会有所不同,请告诉我那个一定会有所不同的地方0(答案:世界上会多一个人)。比较的结果表明:相对于百科知识问题的解答而言,高视觉化的问题与低视觉化的问题一样,都包含上述的视觉空间信息加工网络的活动,而它们的差别在于,比之于低视觉化的问题,高视觉化的顿悟问题的解
决激活了位于左侧额叶的语言区以及双侧的舌回(lingualgyrus)和纺锤状回(fusiformgyrus)(图2B),这几个区域的活动可能与在高视觉化条件下,被试将对问题情境的语言描述转化为视觉表征,并在问题解决的全过程中保持这种表征的活跃状态有关系。
在解决顿悟问题所激活的视觉空间信息加工网络中,楔前叶(precuneus)与心理旋转有关系[39~
44]
,
4 参与思维定势打破的脑机制
有关顿悟的诸种理论的一个共同点,是认为人们在解决顿悟问题时会遇到一种障碍,只有当这种障碍被有效地克服的时候,顿悟问题才会迎刃而解。这种障碍,就是通常所谓的思维定势。比如,在/六火柴问题0中,要求被试用六根火柴构成四个等边三角形。这个问题之所以难,是因为人们一般会假设构成的等边三角形都在一个桌面或者两维的平面上,这就是思维定势。只有当人们打破了这个不正确的思维定势,在一个三维空间搭构火柴时,问题才能有效地被解决。在这方面,一个被良好地界定并
双侧的后部颞中回以及枕中回与动态的视觉有关(比如给被试呈现一个处于下落状态的茶杯相对于给他们呈现放在桌子上的茶杯),而海马旁回与空间定向有关。在顿悟中,人们不但需要转换思考的对象,而且更重要的,他们还要转换思考的背景,正如在心理旋转任务中,人们必须在作为信息加工对象的图形以及图形所处的参照框架之间进行操作和比较一样,这个过程的完成有赖于视觉空间信息加工网络。
226心 理 学 报36卷
被较为充分地研究的问题是Duncker的/蜡烛问题0。在/蜡烛问题0中,被试被要求利用手头的一盒图钉,火柴以及蜡烛等物将蜡烛点燃后固定在木质墙壁上。正确的做法是将图钉盒倒空,用图钉将之固定在墙壁当作烛台(问题的困难之处在于把图钉盒用作烛台)。
我们认为,以/蜡烛问题0为代表的顿悟性问题情境通常包含两个基本的方面,其一,它们包含一种不能够用常规方式顺利解决的问题情境,以/蜡烛问题0为例,通常被试会试图用熔化的蜡液将蜡烛粘在墙上,或者用图钉去固定蜡烛,而这些方法都不能奏效;其二,它包含一个关键性的物体,但这个物体的显而易见的功能却使人们不能够从另一个角度来看待它,比如,图钉盒的显而易见的功能是放置图钉,这种功能是如此/强势0以至于人们往往会忽略它的其他可能的/弱势0用途。抛开一个物体或者概念的/强势0功能或者特性,转而利用其/弱势0功能或者特性,就是一个思维定势的打破过程。为了在实验室条件下模拟这种情形,我们采用了/啊哈谜语0作为实验材料。/啊哈谜语0是Auble等使用的一种谜语材料[45],其特点是用一个含有因果关系的短句意指一种关键性的情景,例如,谜语/因为布破了,所以草垛变得至关重要0意指在跳伞过程中降落伞的布撕裂,跳伞者必须以草垛作为缓冲;谜语/因为是一位专业人士替这位老人照的相,所以看不出照的是谁0意指X光片等。与经典的顿悟问题相类似,上述的谜语情景也包含两个关键性的方面,其一,谜面中所包含的因果关系在通常意义上无法理解;其二,谜语涉及一个关键性概念,这个概念有一个主导的或者/强势0的含义,它压制了其它可能的解释。比如在上述的两个例子中,/布0的基本含义是可穿之物或者用于遮体之物,它极少被用于指称降落伞的面料,而/照片0通常是指人像的写真或摄影,X光片是一种极不典型的照片。在被试对谜语不能达成正确解释的情况下,向他们提供有关答案的关键性提示,比如/降落伞0或者/X光片0,就可以在一瞬间促成他们抛开谜语中那个关键性概念的/强势0含义,转而意识到并且去利用那个概念的/弱势0含义,从而打破思维定势。在实验中,我们还采用了另外一类项目作为基线或者参照活动的状态。这类项目比较容易,人们无须看答案凭自己的能力就能解决,比如/因为太阳光被聚焦,所以报纸就燃烧了起来0(放
*
大镜)或者/因为白色的粉末放进咖啡里,所以咖啡变得甜了0(白糖)。对于这类谜语,人们在看到答案以前就已经有了自己的解答,因此,他们在看到答案时并不会有意外的发现,也就无所谓思维定势的打破。采用事件相关fMRI技术,我们将包含以及不包含思维定势打破的两类心理事件锁定在标准答案呈现的那个时刻点上*。研究的结果表明:包含思维定势打破的项目的平均反应时间为1.87秒(SD=0.52),不包含思维定势打破的项目的平均反应时间为1.09秒(SD=0.17),前者显著长于后者(t(=11)5.98,p<0.001),对于57.8%的包含思维定势打破的项目,被试报告说他们在看到标准答案的瞬间有一种/啊哈0式的发现感;对于75.7%的包含思维定势打破的项目,被试认为答案与谜面匹配良好且十分合理;对于72.9%的包含思维定势打破的项目,被试将他们不能解决问题的主要原因归结为/想到另外一个方面去了0(这一观察从内省的层面证实了这类心理事件的确包含一个思维定势的打破过程)。脑成像数据分析的结果显示:相对于不包含思维定势打破的项目,包含思维定势打破的项目所激活的部位主要有两个,一是扣带前回(布罗德曼24区和32区);二是位于或接近左腹侧额叶的岛叶(布罗德曼13区)和额下回(布罗德曼9区)(图3)。 扣带前回(ACC,anteriorcingulategyrus)位于大脑额叶的内侧面,在解剖上是属于边缘系统的一个部分。认知神经科学的最新研究揭示:在ACC中有一种大型的梭状细胞,这种细胞只有人类以及人类的最近亲)))类人猿才有。这种大型的梭状细胞与大脑皮质的各个部分具有广泛的联系,可能起到调节和改变大脑活动状态的作用(综述见[46]。Vogt等人将ACC的功能概括性地总结为/执行功能0(ex-ecutivefunction)[47]。目前,关于ACC的功能的一个最为普遍的看法是认为ACC参与认知冲突的解决。早先的事件相关电位(ERP)研究表明,在要求被试尽快地作出反应的条件下,有一种错误相关负波(ERN,error-relatednegativity)在人们作出了一个错误反应之后的100到150毫秒之间被观察到,而偶极子分析表明:ERN有一个位于额叶内侧的波源,可能在ACC,这使得人们认为ACC的功能在于监控和补偿错误行为。具体的讲,ACC被假设参与一个比较的过程,它在人们心里想做的正确的反应与其真实做出的反应之间进行比较[48,49]。但是,其后的
LuoJ,NikiK,PhillipsS.Theoriginsofinsightinpuzzlesolving:theroleofanteriorcingulatecortexinbreakingmentalset.Submittedmanuscripts.
2期罗劲:顿悟的大脑机制 227
事件相关fMRI实验却表明:ACC不仅仅在错误反应之后活跃,事实上,只要有反应的冲突,无论其行为上的对错,ACC都会活跃[50]。Carter等人采用由/线索-探测项目0组成的连续操作任务(AX-CPT,ContinuousPerformanceTest),比如A(线索项目))X(探测项目),A)Y,B)X,B)Y等。要求被试只有在探测项目为X而线索项目为A的情况下做反应,对其它情况则不作反应,A)X类项目占到全部项目
数的70%,而其它各类项目只各占10%。A)Y项目和B)X项目所引起的反应冲突会比A)X项目和B)Y项目更多,因为A)Y项目和B)X项目中的A与X都与正性反应相关联。研究结果表明:对于高冲突项目,即使被试作出了正确的选择,ACC也会有较高水平的活动[50]。除此之外,ACC的活动也在Stroop任务中被广泛地观察到[51~53]。
和ACC相类似,左腹侧额叶(LVPFC,leftventro-lateralprefrontalcortex)的功能也与认知冲突以及无关刺激的抑制有关。早期的脑成像研究在语义加工任务中经常观察到LVPFC的活动[54~57],因此,LVPFC曾经一度被认为支持或者参与语义信息的提取。但是,其后的研究却认为LVPFC实际上参与不同语义信息之间的选择。Thompson-Schill等人令被试根据呈现给他们的工具的图片说出与此工具相关联的动词,在低冲突条件下的工具只与少数动词相关,比如剪子只与/剪0相关联;而在高冲突条件下的工具往往与很多动词相关,比如绳子与/系0、/捆0、/扎0、/吊0等很多动词相关联。因此,理论上假设人们在对高冲突条件下的工具作反应时会有较多的语义选择上的冲突与竞争,因为他们必须在众多可能的动词里选择一个(行为反应指标也支持了这个假设)。脑成像研究的结果表明:相对于低冲突条件的项目而言,高冲突条件的项目伴随有明显的LVPFC的活动。在我们所采用的三词判断实验中
[27]
[58]
上面的词有关时(如上面的词为/土地0,下面的词为/钟表0和/音乐0[0关系],或者如上面的词为/土地0,下面的词为/建筑0和/田野0[2关系]),语义冲突与选择较多,因而较之只有一个下面的词与上面的词相关的情形(如上面的词为/土地0,下面的词为/建筑0和/音乐0[1关系])伴随有较多的LVPFC的活动。除参与语义信息的冲突与选择之外,LVPFC还参与工作记忆中的冲突与抑制,在Jonides等的脑成像实验中[59],要求被试在记住一张含有四个英文辅音字母的卡片之后,判断随后出现的探测刺激是否是学过的四个英文字母中的一个,关键性地,他们比较了两类不同的负性(即引起/否0判断的)探测字母所引起的脑活动。其中,/重复0的负性探测字母,虽然它并没有在刚刚学过的卡片中出现,但却在上一轮的/学习-测验0单元中出现过,因此,被试在对它作否定判断时不得不压制对它的熟悉感,从而导致一个抑制过程。而/控制0的负性探测字母则并未在临近的/学习-测验0单元中出现,因此,被试在对,
228心 理 学 报36卷
通过比较/重复0的负性探测字母与/控制0的负性探测字母,Jonides等人确定了参与抑制的关键性脑区在LVPFC(相关的研究见[60])。我们采用汉字材料,比如,在工作记忆任务中令被试学习过/指0字之后,或用与此字音型相似的/脂0字作为探测项目(音型冲突)、或用与此字意义相关的/手0字作为探测项目(语义冲突)、或用一个无关字/全0作为探测项目,结果发现,与最后一种情形相比较,前面的两种条件伴随有明显的LVPFC的活动。Petrides将LVPFC的功能概括为对保持在工作记忆中的信息进行比较和挑选[62]。
根据上述证据,我们认为,相对于不包含思维定势打破的项目,包含思维定势打破的项目所激活的ACC与LVPFC与思维定势打破过程中所产生的认知冲突有关,是思维定势打破过程中的认知冲突导致了这两个区域的活动。
尽管ACC与LVPFC都参与认知冲突的协调,但它们的作用却并不相同。Smith和Jonides(1999)认为:ACC参与较为初级的/前编码反应0(prepro-grammedresponses)的抑制,而LVPFC则参与较为次级的信息加工过程早期的注意与抑制[63]。比如,ACC参与Stroop任务,而在Stroop任务中,信息加工的冲突来源于一个颜色字(比如/蓝0)的字义与这个字本身的颜色(比如用红墨水写的/蓝0字),这种冲突发生在信息加工的初级阶段,换言之,对字义以及字的颜色的加工是人们的固有知识的一部分,是被写进一个/硬性0的程序之中的;而LVPFC参与工作记忆水平的抑制,比如在学习过/指0字以后对/脂0作判断,此时,信息加工的冲突来源于工作记忆的/内存0,亦即:人们对于/指0字的存储是暂时写在一个/软性0的程序之中的。这个差别就好像计算机中的/MicrosoftWord0软件,普通的用户无权修改这个软件本身的某些程序,他们只能建立、修改和存储利用这个软件所作成的文件,在这里,软件本身的程序特性是/硬性0的,它相当于/前编码0阶段的信息加工,而利用这个软件所作的文件则相对是/软性0的,它相当于较为次级的工作记忆与注意水平的信息加工。上述差别的关键在于,它假设ACC参与的是那些被试的自主控制的加工策略所无法左右的信息加工过程,而LVPFC则参与的是那些被试可以自主控制的信息加工过程。换言之,ACC的作用在于调节那些预料之外的、突发性的认知冲突;而左腹侧额叶的作用,则在于调节那些预料之内的、常规的认知冲[61]
目前,有两个方面的证据支持上述的设想。其
一,在Stroop任务中,当被试的加工策略性控制较高时,ACC的活动水平较低;而当被试的加工策略性控制较低时,ACC的活动水平较高[64]。在Carter等人的这项研究中,他们改变了在一个区组中/一致性项目0(用红墨水写的/红0字)与/不一致性项目0的比例。理论上认为,如果/不一致性项目0在一个区组的全部项目中所占的比例越高,被试的警觉性就越高,因此他们的加工策略性控制就越强;而/一致性项目0所占的比例越高,被试的警觉性就越低,因此他们的加工策略性控制就越弱。研究的结果表明:在/不一致性项目0所占的比例高的情况下,被试对/不一致性项目0的反应时较短,而ACC的活动水平较低;而在/一致性项目0比例高的情况下,被试对/不一致性项目0的反应时较长,而ACC的活动水平较强。这说明了ACC的作用在于调节那些预料之外的、突发性的认知冲突。其二,威斯康星卡片分类任务(WCST,WisconsinCardSortingTask)只需要LVPFC的参与,不需要ACC的参与。WCST要求被试根据图形的数量、或者形状、或者颜色的维度来对卡片进行分类。但是分类的维度是由主试控制而随时变换的,被试并不知道这种变换何时会发生,因此,他们只能在得到反馈时调整分类的方式。在WCST中也包含着思维定势的改变,但这种改变有别于顿悟中所发生的改变,因为在WCST中,被试明确地知道他将根据图形的数量/形状/颜色来分类,并不会有别的可能性,所以,在WCST中思维定势的改变只能算作一种/转移0而非/打破0。Monchi等人对于WCST的事件相关fMRI研究表明[65]:相对于收到正反馈而言,当被试收到负反馈并不得不改变其分类策略时,LVPFC就活跃。尽管与基线水平相比较,当被试收到负反馈时也会有ACC的活动,但ACC活动的水平与收到正反馈时一样强,这说明ACC在其中的作用仅是无特异性的注意性控制加工而已,并非WCST中思维定势/转移0的关键神经结构。
先前的认知心理学实验表明:顿悟问题的解决是在被试的认知加工策略控制范围之外发生的。Metcalfe的一系列研究证明,与普通推理问题的解决有所不同,顿悟问题的解决在元认知监控(FOK,fee-ling-of-knowing)的范围之外,换言之,人们不能监控顿悟问题的解决[32~34]。基于以上的证据,我们提出了一个有关思维定势改变的一般假设,即:对于
2期罗劲:顿悟的大脑机制 229
的/转移0(比如WCST)而言,有LVPFC的参与就足够了;但对于在人们的加工策略的控制范围之外的思维定势的/打破0(比如顿悟)而言,则需要ACC与LVPFC的协同参与。上述假设包含如下的理论预期:一,它预期ACC活跃于思维定势打破的早期阶段,起到一个/早期预警系统0(earlywarningsystem)的作用;二,它预期当问题解决者了解了顿悟问题的结构或者经过反复练习摸清了顿悟问题的构成规律并且发展出一般性的信息加工控制策略的时候,ACC的活动会降低。目前,我们已经获得了四个证据支持上述预测:
第一,对于包含思维定势打破的项目而言,项目理解的难易程度只会影响LVPFC的活动,但并不影响ACC的活动。我们比较了两类包含思维定势打破的项目(即谜语的答案),对于一类谜语的答案,被试评价比较容易理解,认为其意义显而易见;而对于另一类谜语的答案,被试认为可以理解,但是颇费周折。相对于不包含思维定势打破的项目而言,无论是较难的还是较易的包含思维定势打破的项目,都伴随有ACC和LVPFC的活动;但是,相对于较易的包含思维定势打破的项目而言,较难项目的反应时较长,而且伴随有更多的LVPFC的活动。这个结果说明:只有LVPFC参与了较难的答案的理解,ACC并未参与其中;换言之,只有LVPFC对于任务的难度有反应。这一结果与ACC活跃于思维定势打破的早期阶段的设想是一致的。在被试看到标准答案的一瞬间,原有的思维定势被摆脱,这是思维定势的/打破0过程,而在被试确定了正确的思路之后,他们沿着这条思路在几种可能的解释之间加以选择和评价,并最终达成理解,这是思维定势的/转移0过程。相对于不包含思维定势打破的项目而言,无论是较难的还是较易的包含思维定势打破的项目都伴随有ACC的活动,这说明了两类项目都包含有/打破0过程,而较难的项目与较易的项目之间的差别是/后打破性0(post-breaking)的,它们发生于思维定势的/转移0阶段,因而较难的项目与较易的项目之间的差别就只存在于LVPFC。
第二,ERP技术可以记录大脑进行信息加工时的电活动,因而有可能精确记录顿悟过程的脑活动的时间特征。我们做了一个与上述的fMRI实验平行的ERP实验,实验的结果表明:与不包含思维定势打破的项目/答案相比较,包含思维定势打破的
*
项目/答案显示一个波峰为380毫秒的差异负波(N380)。从目前掌握的资料来看,对这个N380,有三种可能的解释,其一,实验所观察到的N380是N2,N2成分在EriksenFlanker任务中被观察到[67],它反映认知加工的冲突,其反应的潜伏期为340~380毫秒,其波源在ACC;其二,实验所观察到的N380是ERN[48,49]。ERN在人们作出了一个错误反应之后的100到150毫秒之间被观察到,其波源也在ACC,ERN反映了错误行为的监控和补偿。其三,实验所观察到的N380是N400[68],N400在人们加工与一个句子的语境不相符合的信息时被观察到(比如,对于句子/他在咖啡里加入了)))0,/糖0是符合语境预期的信息,而/袜子0则不是),一般认为,N400的波源在颞叶[69~
71]
。根据我们在实验中所观
察到的N380的强度分布状况以及其他特征,我们初步认为N380是N2或者ERN,而不是N400。我们进一步利用偶极子分析方法研究了N380的波源,结果证实了原先的设想,即在顿悟中观察到的N380的波源在ACC。考虑到整个的顿悟过程从启动到完成需要约2000毫秒左右的时间,因此,ACC的活动是发生在顿悟过程启动的初级阶段,在思维定势的打破中起到一个/早期预警系统0的作用。
第三,我们的假设预期:当问题解决者了解了顿悟性问题的结构或者经过反复练习摸清了顿悟性问题的构成规律并且发展出一般性的信息加工控制策略的时候,ACC的活动会降低。为了验证这个假设,我们比较了两类不同的谜语,对于这两类谜语,我们都通过了一个事先的筛选过程,保留那些被试能够很好地理解其谜面、但却并不知道答案的项目,并在fMRI扫描时给其提供标准答案。实验的关键设计是:对于其中的一组谜语,被试知道其构成方式,而对于另外一组谜语,被试不知道其构成方式。对于前者,我们采用了日本传统的/同音谜语0(homo-phoneriddles),这类谜语的特点是用同音字法构成谜语,比如/横纲若乃花(最著名的相扑士)去神社祈祷,但却只获得15战12胜的赛绩,为什么?0答案是/参拜0(日语中/参拜0与/三败0同音),再比如/屋子的什么地方人们去了会变得衰老?0答案是/廊下0(/廊下0与/老化0同音)等。而对于后者,我们采用脑筋急转弯问题作为材料,但由于这类问题千变万化,所以被试并不知道谜语的结构。结果表明,相对于静息水平而言,LVPFC在两类谜语的解决中的参
*
[66]
LuoJ,NikiK.Unpublishedobservations.
230心 理 学 报36卷
与程度相当,其阈限以上的兴奋的体积均为1000个像素左右(voxel,1个像素相当于2mm@2mm@2mm的体积;/同音谜语0:1072voxel;/脑筋急转弯问题0:1262voxel),但是ACC在/脑的体操谜语0的解决中的活动却明显高于/同音谜语0的解决(/同音谜语0:24voxel;/脑筋急转弯问题0:912voxel)。这说明:如果在猜谜者知道谜语的构成方式的情况下,解谜过程主要依靠LVPFC而不是ACC。
第四,在上述以/啊哈谜语0作为实验材料的实验中,我们将每个被试解决谜语的全部过程均等地分为早期的三分之一、中期的三分之一和后期的三分之一三个阶段/区组,并且分别在这三个阶段之间进行了比较,结果发现,相对于安静休息的参照状态而言,早、中、后三个阶段都伴随有明显的ACC的活动。但是,相对于中期和后期两个阶段而言,早期阶段所激发的ACC的活动明显地要多。这种差异不是由于在早、中、后三个阶段中的谜语项目的性质或者组成不同所造成的,事实上,这三个阶段中所包含的思维定势打破的项目的比例相当,且全部项目的呈现顺序在各个不同的被试是随机的。我们认为,虽然在正式的实验之前,被试作了足够数量的谜语练习,但仍有可能在该类谜语解决的早期(前三分之一)阶段,被试对谜语的构成规律所知相对较少,因而没有办法采取一般的信息加工策略来控制局面,所以每当谜语的答案呈现,就需要ACC来控制最初的信息加工的流向。但是,随着所做项目的增加,被试逐渐有意识或者无意识地获得了一些有关谜语构成规律的知识,这使得他们有能力采取一定的信息加工策略来控制局面,从而降低了对于ACC的依赖。但这并不是说,由于练习的作用使得被试获得了某种问题解决的技能,因为目前已知的对于认知技能的获得较为敏感的区域,比如前部的前额叶和背外侧的额叶[72],在上述的三个阶段中的参与程度并无明显差别。上述的四点观察为我们有关思维定势打破的脑神经过程的假设提供了一定的证据。 Kounios等人根据其最新的ERP研究结果,认为ACC的活动与人们在问题解决中所遇到的/障碍0(block)有关[73],但我们却认为:ACC的活动与/障碍0状态无关,而与/障碍的解决0过程有关。/障碍0状态是指被试在问题解决过程中遇到明显的困难,处于一种不知所措的状态;而/障碍的解决0过程则是指被试成功地找到了能有效地解决问题的新途径。从理论上讲,/障碍0状态是一种思维的/停顿0/发生的ACC。为了验证上述假设,我们进一步比较了被试在加工两种类型的句子时的大脑活动,一类是被试(无须看答案就)能够理解简单的/啊哈谜语0(比如/因为太阳光被聚焦,所以报纸就燃烧了起来0)))放大镜),另一类是被试(不看答案就)不能理解的困难的/啊哈谜语0(比如/因为是一位专业人士替这位老人照的相,所以看不出照的是谁0)))X光片)。由于被试在加工困难的/啊哈谜语0时,经历了/障碍0状态,因此,我们预期,如果ACC参与/障碍0状态的话,那么,ACC在被试读那些不能理解的困难的句子时的活动将会更加强烈。反之,如果ACC参与/障碍的解决0过程的话,那么它将在被试读那些可以理解的简单的句子时的活动得更多(即使是简单的/啊哈谜语0,亦包含一种间接含糊的表达,因而,被试在阅读这类句子时也需要克服某种障碍)))尽管这种障碍没有困难的/啊哈谜语0中的障碍强大)。研究结果表明:在谜面的呈现阶段,不能理解的困难的/啊哈谜语0相对于能够理解简单的/啊哈谜语0激活左侧的颞中回以及右侧的额中回;而能够理解简单的/啊哈谜语0相对于不能理解的困难的/啊哈谜语0则激活ACC以及额叶内侧面的区域。这说明ACC的活动与/障碍0状态无关,而与/障碍的解决0过程有关。
5 顿悟与创造性思维
研究顿悟的大脑机制的更为一般性的目的,是探讨创造性思维的本质。进入知识经济时代以后,创造性思维变得非常重要。顿悟是创造性思维的一个重要的心理基础,对顿悟的研究,可以澄清目前创造性思维理论中的许多含糊不清的基本问题。
目前,有关创造型思维的脑过程的设想主要涉及左右大脑半球的功能区分,认为左侧大脑参与常规性的思维,而右侧大脑则参与创造性的思维。其证据主要来自脑损伤病人以及一些视觉呈现技术(比如将视觉刺激呈现到单侧的视野上)[74~76]。但显而易见,上述方法过于粗略而无法使探索变得深入和具体。脑成像技术可以以很高的空间分辨率记录大脑在瞬间的信息加工活动,因而有可能为我们提供更加详细的有关顿悟的脑活动状况,并将研究的焦点转移至更加精确的脑结构上。例如,有研究表明,海马在人做梦的时候会活动,而亦有经验的观察表明,梦境与创造性思维有关;这样,就有可能意味着海马的活动与创造性思维有关。利用脑成,[77]
2期
*
罗劲:顿悟的大脑机制 231
现实生活情景的回忆的大脑区域,一个主要的发现是,相对于现实生活情景的回忆,梦境的回忆会激活海马与左侧额叶,考虑到该实验结果是在小心地控制了那些已知有可能导致海马活动的无关的变量)))特别是记忆时程的长短、记忆内容的空间特征
[78]
以及记忆细节的丰富性等)))的情况下获得的,因此我们认为海马在回忆梦境中的活动可能与梦境中那些奇异的想象有关系,而按照精神分析的看法,这些想象则可能源于梦者内心的某种需要。
关于柯勒的顿悟的一个更为广泛的思想背景是德国的理性主义传统。在与柯勒同时代的德国学者中,还有另外一位从一个更加广泛的人类学和宗教学的角度提出了类似的理念,这个人就是对后世产生广泛影响的德国哲学家卡西尔(ErnstCassir-er)[79]。就像柯勒反对当时占主导地位的桑代克的尝试-错误理论一样,卡西尔反对传统的语言和概念的产生理论。传统理论认为:原始语言或者概念的形成过程,是一个由特殊到普遍的过程,即一个抽象的过程,比如,人们在品食了各种水果之后,就会逐渐抛弃其特殊性,保留其普遍性,从而形成一个关于/水果0的概念。卡西尔认为,这种假设是不合理的,因为抽象乃是语言的功能,而在语言尚未产生的条件下,人们并不具有这种能力。卡西尔将原始语言的产生与原始神话的产生联系起来,假设了一个由/伴随强烈需要的满足而产生的高度注意状态0,到/瞬息神(momentarydeities)的崇拜0,到/原始概念的形成0的过程。这一过程的核心认为,原始人会对那些极大地满足了他们的需要的事物产生原始宗教式的崇拜,这种崇拜以高度的注意集中为特征并把瞬息万变的/意识之流0固化在一个点上,而这种固化的意识形态,就是原始概念的雏形。卡西尔的这一假设的主要证据,来源于有关原始部落瞬息神崇拜的人类学研究。这些研究显示:原始部落的人会把那些极大的满足过自己的需要的事物遵奉为瞬息神,比如,一眼在当事人极度干渴时发掘出来的泉水,一个曾经在猛兽出现的时刻供其藏身的山洞,或者一棵他们在其簌簌的枝叶之下突然感觉敬畏的树等。这种崇拜与其后才发展起来多神崇拜(比如佛教)与到再后来才发展起来的统一神崇拜(比如基督教)有所不同,是宗教崇拜的最原始的形式,因其往往源于一种瞬间的心理体验,故而被称为瞬息神。卡西尔设想的这一发现过程与柯勒的顿悟过程的共
*
同之处在于:第一,它们都假设了一个瞬间的、意识高度集中的状态;第二,它们都假设了在这个状态中,人们的生理的或心理的需要被满足,亦即任务相关性;第三,它们都假设人们在这个状态中所获得的意识与体验被深刻地保存在记忆之中,并对其以后的认知与行为产生深远的影响(一个例子是,那句名言/我找到了!0(Eureka或0Ihavefoundit0)有两重的意指,一是指阿基米德对皇冠问题的顿悟性解决,二是指美国早期的淘金者发现金矿)。而目前我们已经知道,海马的功能至少与其中的两个特征有关,一是它在任务相关性联系的形成过程中起反应,二是它是支持长时记忆的关键的脑结构。
将/卡西尔过程0与顿悟联系起来的理论意义在于,它将顿悟置于一个更为广泛的社会历史背景之下,使顿悟有可能延伸到概念形成甚至宗教心理学的领域。例如,目前认知心理学的概念形成的研究主要集中在讨论人们如何通过一个抽象过程,逐步将不同的个例的一般特征抽取出来,并最终形成一个普遍适用的概念。而顿悟理论则假设了一个相反的过程,它假设概念的形成是一个突发的过程,人们透过个例,在瞬间窥见事物的某种本质联系,并逐步将这种领悟推广于其他事物)))这是一个类似于科学规律的发现与推广过程,这个过程强调领悟与概念形成的突然发生的特性与一蹴而就的特性。
6 小结
尽管目前对于顿悟的大脑机制的研究还处于起步阶段,但是利用脑成像技术以及谜语材料我们业已获得了一个粗略的有关顿悟的脑过程框架,这个框架假设顿悟过程是由作为早期预警系统的ACC所发动,并由负责新异有效联系(任务相关联系)形成的海马、负责思维定势转换和语言加工的LVPFC以及负责思考的背景或参照框架切换的视觉空间信息加工网络协同完成的。
参
考
文
献
1 EpsteinR,etal.0Insight0inthepigeon:antecedentsanddeterminants
ofanintelligentperformance.Nature,1984,308:61~62
2 SchoolerJW,OhlssonS,BrooksK.Thoughtsbeyondwords:when
languageovershadowsinsight.JournalofExperimentalpsychology:Gen-eral,1993,122:166~183
3 SternbergRJ,DavidsonJE.Thenatureofinsight.Cambridge,Mas-sachusetts:theMITPress,1995
LuoJ,NikiK.Unpublishedobservations.
232心 理 学 报36卷
4 KohlerW.Thementalityofapes.London:Routledge&KeganPaul,
1925
5 WertheimerM.Productivethinking.NewYork:Harper,19596 KnoblichG,OhlssonS,HaiderH,etal.Constraintrelaxationand
chunkdecompositionininsightproblemsolving.JournalofExperimentalPsychology:Learning,MemoryandCognition,1999,25:1534~15567 KnoblichG,OhlssonS,RaneyGE.Aneyemovementstudyofinsight
problemsolving.MemoryandCognition,2001,29:1000~10098 OhlssonS.Restructuringrevisited:IIAninformationprocessingtheory
ofrestructuringandinsight.ScandinavianJournalofPsychology,1984,25:117~129
9 OhlssonS.Information-processingexplanationsofinsightandrelated
phenomena.In:GilhooleyKJ.AdvancesinthePsychologyofThink-ing.London:Harvester-Wheatsheaf,1992.1~44
10 KeaneM.ModelingproblemsolvinginGestalt0insight0problems.Irish
JournalofPsychology,1989,10:201~215
11 SmithSM.Gettingintoandoutofmentalruts:Atheoryoffixation,in-cubation,andinsight.InSternbergRJ,DavidsonJE.Thenatureofin-sight.Cambridge,Massachusetts:theMITPress,1995,229~25112 LuchinsAS,LuchinsEH.Newexperimentalattemptsatpreventing
mechanizationinproblemsolving.JournalofGeneralPsychology,1950,42:279~297
13 MacGregorJN,OrmerodTC,ChronicleEP.Informationprocessing
andinsight:aprocessmodelofperformanceonthenine-dotandrelat-edproblems.JournalofExperimentalPsychology:Learning,MemoryandCognition,2001,27:176~201
14 WeisbergRW.Creativity:Geniusandothermyths.NewYork:Free-man,1986.50
15 WindholzG.Pavlovvs.Kohler.Pavlov.slittle-knownprimatere-search.PavlovJBiolSci,1984,19:23~31
16 LuoJ,NikiK.Functionofhippocampusin0insight0ofproblemsolv-ing.Hippocampus,2003,13:316~23
17 EichenbaumH,OttoT,CohenNJ.Twofunctionalcomponentsofthe
hippocampalmemorysystem.BehavioralandBrainSciences,1994,17:499~518
18 CohenNJetal.Hippocampalsystemanddeclarative(relational)mem-ory:summarizingthedatafromfunctionalneuroimagingstudies.Hip-pocampus,1999,9:83~98
19 McClellandJL,McNaughtonBL,O.ReillyRC.Whytherearecom-plementarylearningsystemsinthehippocampusandneocortex:insightsfromthesuccessesandfailuresofconnectionistmodelsoflearningandmemory.PsychologyReview,1995,102:419~457
20 CorkinS.H.M..sdetectionanddescriptionofambiguousmeanings.
Unpublishedtranscript,1973,Dept.ofBrainandCognitionSciences,M.I.T.,Cambridge,MA.In:MacKayDG,StewartR,BurkeDM.H.M.revisited:relationsbetweenlanguagecomprehension,memory,andthehippocampalsystem.JournalofCognitiveNeuroscience,1998,10:377~394
21 TulvingE,MarkowitschHJ,KapurS,etal.Noveltyencodingnet-worksinthehumanbrain:positronemissiontomographydata.NeuroRe-port,1994,5:2525~2528
22 HJ,Craiketal.tyandactivationsinPETstudiesofmemoryencodingandretrieval.CerebralCortex,1996,6:71~79
23 KnightR.Contributionofhumanhippocampalregiontonoveltydetec-tion.Nature,1996,383:256~259
24 GrunwaldT,LehnertzK,HeinzeHJ,etal.Verbalnoveltydetection
withinthehumanhippocampusproper.ProceedingsoftheNationalA-cademyofSciencesoftheUnitedStatesofAmerica,1998,95:3193~3197
25 RedishAD.Thehippocampaldebate:areweaskingtherightques-tions?BehaviouralBrainResearch,2001,127:81~98
26 EichenbaumH,etal.Cue-samplingandgoal-approachcorrelatesof
hippocampalunitactivityinratsperforminganodor-discriminationtask.TheJournalofNeuroscience,1987,7:716~732
27 LuoJ,NikiK.Roleofmedialtemporallobeinextensiveretrievalof
task-relatedknowledge.Hippocampus,2002,12:487~49428 HenkeK,etal.Humanhippocampusassociatesinformationinmemory.
ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1999,96:5884~5889
29 HartleyT,etal.Thewell-wornrouteandthepathlesstraveled:dis-tinctneuralbasesofroutefollowingandwayfindinginhumans.Neuron,2003,37:877~888
30 SimonH.Thescienceoftheartificial.Cambridge,MA:MITPress,
1969.153
31 DurkinHE.Trial-and-error,gradualanalysisandsuddenreorgan-i
zation.Anexperimentalstudyofproblemsolving.Archpsychol,1937,210:1~85
32 MetcalfeJ.Feelingofknowinginmemoryandproblemsolving.Journal
ofExperimentalPsychology:Learning,Memory,andCognition,1986,12:288~294
33 MetcalfeJ.Premonitionsofinsightpredictimpendingerror.Journalof
ExperimentalPsychology:Learning,Memory,andCognition,1986,12:623~634
34 MetcalfeJ,WiebeD.Intuitionininsightandnoninsightproblemsolv-ing.MemoryandCognition,1987,15:238~246
35 MaierNRF.ReasoninginhumansII.Thesolutionofaproblemandits
appearanceinconsciousness.JournalofComparativePsychology,1931,12:181~194
36 SmithRW,KouniosJ.Suddeninsight:all-or-noneprocessingre-vealedbyspeed-accuracydecomposition.JournalofExperimentalPsy-chology:Learning,Memory,andCognition,1996,22:1443~146237 GoelV,BuchelC,FrithC,etal.Dissociationofmechanismsunderly-ingsyllogisticreasoning.NeuroImage,2000,12:504~514
38 GoelV,DolanRJ.Functionalneuroanatomyofthree-termrelational
reasoning.Neuropsychologia,2001,39:901~909
39 HarrisIM,MiniussiC.ParietalLobeContributiontoMentalRotation
DemonstratedwithrTMS.JournalofCognitiveNeuroscience,2003,15:315~323
40 JordanK,HeinzeHJ,LutzK,etal.Corticalactivationsduringthe
mentalrotationofdifferentvisualobjects.NeuroImage,2001,13:143~152
41 VingerhoetsG,SantensP,VanLaereK,etal.Regionalbrainactivity
ofmentalinvol:
2期罗劲:顿悟的大脑机制
America,1997,94:14792~14797
233
positronemissiontomographystudy.NeuroImage,2001,13:381~39142 HarrisIM,EganGF,SonkkilaC,etal.Selectiverightparietallobe
activationduringmentalrotation:aparametricPETstudy.Brain,2000,123:65~73
43 CarpenterPA,JustMA,KellerTA,etal.Gradedfunctionalactiva-tioninthevisuospatialsystemwiththeamountoftaskdemand.JournalofCognitiveNeuroscience,1999,11:9~24
44 KosslynSM,DiGirolamoGJ,ThompsonWL,etal.Mentalrotationof
objectsversushands:neuralmechanismsrevealedbypositronemissiontomography.Psychophysiology,1998,35:151~161
45 AublePM,FranksJJ,SoraciSA.Effordtowardcomprehension:Elab-orationor0aha0?MemoryandCognition,1979,7:426~43446 AllmanJM,etal.Theanteriorcingulatecortex.Theevolutionofan
interfacebetweenemotionandcognition.AnnalsoftheNewYorkA-cademyofSciences,2001,935:107~117
47 VogtBA,FinchDM,OlsonCR.Functionalheterogeneityincingu-latecortex:theanteriorexecutiveandposteriorevaluativeregions.CerebralCortex,1992,2:435~443
48 HohnsbeinJ,FalkensteinM,HoormanJ.Errorprocessinginvisualand
auditorychoicereactiontimetasks.JournalofPsychophysiology,1989,3:32
49 GehringW,GossB,ColesMGH,etal.Aneuralsystemforerrorde-tectionandcompensation.PsychologicalScience,1993,4:385~39050 CarterCS,BraverTS,BarchDM,etal.Anteriorcingulatecortex,
errordetection,andtheonlinemonitoringofperformance.Science,1998,280:747~749
51 PardoJV,PardoPJ,JanerKW.Theanteriorcingulatecortexmed-i
atesprocessingselectionintheStroopattentionalconflictparadigm.Pro-ceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1990,87:256~259
52 BenchCJ,etal.Investigationsofthefunctionalanatomyofattention
usingtheStrooptest.Neuropsychologia,1993,31:907~92253 CarterCS,MintunM,CohenJD.Interferenceandfacilitationeffects
duringselectiveattention:anH215OPETstudyofStrooptaskperfor-mance.NeuroImage,1995,2:264~272
54 BucknerRL,PetersenSE,OjemannJG,etal.Functionalanatomical
studiesofexplicitandimplicitmemoryretrievaltasks.TheJournalofNeuroscience,1995,15(1Pt1):12~19
55 DembJB,DesmondJE,WagnerAD,etal.Semanticencodingand
retrievalintheleftinferiorprefrontalcortex:afunctionalMRIstudyoftaskdifficultyandprocessspecificity.TheJournalofNeuroscience,1995,15:5870~5878
56 DemonetJF,CholletF,RamsayS,etal.Theanatomyofphonological
andsemanticprocessinginnormalsubjects.Brain,1992,115:1753~1758
57 KapurS,CraikFI,TulvingE,etal.Neuroanatomicalcorrelatesofen-codinginepisodicmemory:levelsofprocessingeffect.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1994,91:2008~2011
58 Thompson-SchillSL,D.EspositoM,etal.Roleofleftinferiorpre-frontalcortexinretrievalofsemanticknowledge:areevaluation.Pro-ofofof59 JonidesJ,SmithEE,MarshuetzC,etal.Inhibitioninverbalworking
memoryrevealedbybrainactivation.ProceedingsoftheNationalA-cademyofSciencesoftheUnitedStatesofAmerica,1998,95:8410~8413
60 LuoJ,NikiK,DingZG,etal.TheHierarchicalStructureofFrontal
LobeinMediatingthePerceptualLevelInhibitionandWorkingMemoryLevelInhibition.ActaPsychologicaSinica,2003,35:427~432 (罗劲,仁木和久,丁之光等.额叶皮层内知觉干扰与工作记忆
干扰引起的抑制.心理学报,2003,35:427~432)
61 NikiK,LuoJ.Thehierarchicalstructureofprefrontalcortexsubserving
directandindirectinhibitioninrecognition.ProceedingofSeventhIn-ternationalConferenceonNeuralInformationProcessing,2000,862~867
62 PetridesM.Frontallobesandworkingmemory:evidencefrominvestiga-tionsoftheeffectsofcorticalexcisionsinnonhumanprimates.In:BollerF,GrafmanJeds.HandbookofNeuropsychology(Vol9),Ams-terdam:Elsevier,1994,59~81
63 SmithEE,JonidesJ.Storageandexecutiveprocessesinthefrontal
lobes.Science,1999,283:1657~1661
64 CarterCS,MacdonaldAM,BotvinickM,etal.Parsingexecutivepro-cesses:strategicvs.evaluativefunctionsoftheanteriorcingulatecor-tex.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2000,97:1944~1948
65 MonchiO,PetridesM,PetreV,etal.WisconsinCardSortingrevisit-ed:distinctneuralcircuitsparticipatingindifferentstagesofthetask-identifiedbyevent-relatedfunctionalmagneticresonanceimaging.TheJournalofNeuroscience,2001,21:7733~7741
66 MaiX,LuoJ,WuJ,etal.0Aha!0effectsinaguessingriddletask:
anERPstudy.Posterpresentedinthe9thAnnualMeetingoftheOrga-nizationforHumanBrainMapping(HBM2003)
67 VanVeenV,CarterCS.Thetimingofaction-monitoringprocessesin
theanteriorcingulatedcortex.JournalofCognitiveNeuroscience,2002,14:593~602
68 KutasM,HillyardSA.Readingsenselesssentences:Brainpotentials
reflectsemanticincongruity.Science,1980,207:203~205
69 ElgerCE,GrunwaldT,LehnertzK,etal.Humantemporallobepoten-tialsinverballearningandmemoryprocesses.Neuropsychologia,1997,35:657~667
70 McCarthyG,NobreAC,BentinS,etal.Language-relatedfieldpo-tentialsintheanterior-medialtemporallobe.I.Intracranialdistribu-tionandneuralgenerators.TheJournalofNeuroscience,1995,15:1080~1089
71 SmithME,StapletonJM,HalgrenE.Humanmedialtemporallobepo-tentialsevokedinmemoryandlanguagetasks.Electroencephalographyandclinicalneurophysiology,1986,63:145~159
72 PoldrackRA,PrabhakaranV,SegerCA,etal.Striatalactivation
duringacquisitionofacognitiveskill.Neuropsychology,1999,13:564~574
73 KouniosJ,BeemanMJ,LiuS,FrymiareJ,AngelakisE,Stathopoulou
S.TheSparkofInsight:ElectrophysiologicalCorrelatesofthe0AHA!0inpresCognitive-
234
scienceSocietyMeetinginApril2003
心 理 学 报
6:545~573
36卷
74 BeemanMJ,BowdenEM.Therighthemispheremaintainssolution-relatedactivationforyet-to-besolvedinsightproblems.Memory&Cognition,2000,28,1231~1241
75 BowdenEM,BeemanMJ.Gettingtherightidea:Righthemisphere
contributionstosolvinginsightproblems.PsychologicalScience,1998,9:435~440
76 BowdenEM.Theeffectofreportableandunreportablehintsonanagram
solutionandtheaha!experience.ConsciousnessandCognition,1997,
77 LouieK,WilsonMA.Temporallystructuredreplayofawake
hippocampalensembleactivityduringrapideyemovementsleep.Neu-ron,2001,29:145~156
78 NikiK,LuoJ.AnfMRIstudyonthetime-limitedroleofthemedial
temporallobeinlong-termtopographicalautobiographicmemory.Jour-nalofCognitiveNeuroscience,2002,14:500~507
79 CassirerE.LanguageandMyth.NewYork,NY:DoverPublications
Inc,1946
NEURALCORRELATESOFINSIGHT
LuoJing
(KeyLaboratoryofMentalHealth,ChineseAcademyofSciences,Beijing100101,China)
Abstract
SincetheworkofWolfgangKohler,theprocessofinsightinproblemsolvinghasbeenthesubjectofconsiderableinvestiga-tion.Yet,theneuralcorrelatesofinsightremainsunknown.Asthesuddenandunexpectedchangeofone.spointofviewthati-lluminatesashortandelegantsolutionpathtoaparticularproblem,insightmeansformingofnovel,efficientassociationsamongtheoldconceptsandbreakingoftheunwarrantedmentalset.Subjectswereimagedbyfunctionalmagneticresonanceimaging(fMRI)whentheyweresolvingtheriddlesandpuzzles.Resultsshowedactivitiesinfrontal,temporal,andparietalareastobeassociatedwiththeprocessofinsight.Theseresults,togetherwithotherevidences,suggestedthat(1)hippocampusmediatedtheprocessofformingofnovel,efficientassociationsininsight;(2)aspatialneuralnetworkincludingbilateralmiddletemporal/occipitalgyrus,leftmiddlefrontalgyrus,andleftparahippocampalgyrusmediatedtherepresentationalchangeininsight;and(3)anteriorcingulatecortexandleftlateralprefrontalcortexmediatedthebreakingofunwarrantedmentalsetininsight.Keywords insight,even-trelatedfMRI,problemsolving.
探讨顿悟的心理过程与大脑机制
)))评罗劲的5顿悟的大脑机制6
MENTALPROCESSANDBRAINMECHANISMOFINSIGHT:
COMMENTSONLUO.SNEURALCORRELATESOFINSIGHT
傅小兰
(中国科学院心理研究所认知心理学研究室,北京100101)
罗劲的5顿悟的大脑机制6一文,总结了他利用脑成像技术与谜语材料进行的一系列实验研究,并结合已有的文献资料与他本人的理论思考,为我们绘制出顿悟的大脑机制框架
的首幅草图:顿悟过程是由作为早期预警系统的扣带前回(anteriorcingulategyrus,简称ACC)所发动,由负责新异有效联系(任务相关联系)形成的海马(hippocampus)、负责思维定势