病毒是颗粒很小、以纳米为测量单位、结构简单、寄生性严格,以复制进行繁殖的一类非细胞型微生物。病毒是比细菌还小、没有细胞结构、只能在活细胞中增殖的微生物。由蛋白质和核酸组成。多数要用电子显微镜才能观察到。病毒可以利用宿主的细胞系统进行自我复制,但无法独立生长和复制。病毒可以感染所有的具有细胞的生命体。第一个已知的病毒是烟草花叶病毒,由马丁乌斯・贝杰林克于1899年发现并命名,已有超过5000种类型的病毒得到鉴定。研究病毒的科学被称为病毒学,是微生物学的一个分支。
病毒_病毒 -简介
流感病毒
病毒是一类个体微小,结构简单,只含单一核酸(DNA/RNA),必须在活细胞内寄生并以复制方式增殖的非细胞型微生物。病毒同所有生物一样,具有遗传、变异、进化的能力,是一种体积非常微小,结构极其简单的生命形式,病毒有高度的寄生性,完全依赖宿主细胞的能量和代谢系统,获取生命活动所需的物质和能量,离开宿主细胞,它只是一个大化学分子,停止活动,可制成蛋白质结晶,为一个非生命体,遇到宿主细胞它会通过吸附、进入、复制、装配、释放子代病毒而显示典型的生命体特征,所以病毒是介于生物与非生物的一种原始的生命体。
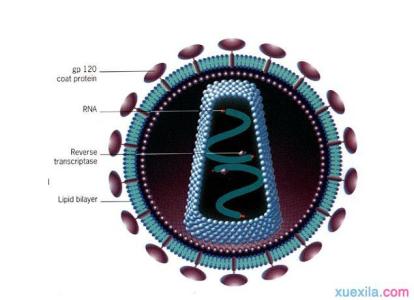
病毒由两到三个成份组成:病毒都含有遗传物质(RNA或DNA,只由蛋白质组成的朊病毒并不属于病毒); 所有的病毒也都有由蛋白质形成的衣壳,用来包裹和保护其中的遗传物质;此外,部分病毒在到达细胞表面时能够形成脂质的包膜环绕在外。病毒的形态各异,从简单的螺旋形和正二十面体形到复合型结构。病毒颗粒大约是细菌大小的百分之一。
基本信息
概述
①形体极其微小,一般都能通过细菌滤器,因此病毒原叫“过滤性病毒”,必须在电子显微镜下才能观察。不同病毒的大小变动于20~450纳米之间。最大的为痘病毒科,大小为(170~260)×(300~450)纳米,最小的为双联病毒科,直径18~20纳米。
②没有细胞构造,其主要成分仅为核酸和蛋白质两种,故又称“分子生物”;
③每一种病毒只含一种核酸,不是DNA就是RNA
④既无产能酶系,也无蛋白质和核酸合成酶系,只能利用宿主活细胞内现成代谢系统合成自身的核酸和蛋白质成分
⑤以核酸和蛋白质等“元件”的装配实现其大量繁殖
⑥在离体条件下,能以无生命的生物大分子状态存在,并长期保持其侵染活力
⑦对一般抗生素不敏感,但对干扰素敏感。
⑧有些病毒的核酸还能整合到宿主的基因组中,并诱发潜伏性感染。
病毒_病毒 -特性
病毒性质的两重性;
一、病毒生命形式的两重性
1.病毒存在的两重性病毒的生命活动很特殊,对细胞有绝对的依存性。其存在形式有二:一是细胞外形式,一是细胞内形式。存在于细胞外环境时,则不显复制活性,但保持感染活性,是病毒体或病毒颗粒形式。进入细胞内则解体释放出核酸分子(DNA或RNA),借细胞内环境的条件以独特的生命活动体系进行复制,是为核酸分子形式。
⒉病毒的结晶性与非结晶性病毒可提纯为结晶体。我们知道结晶体是一个化学概念,是很多无机化合物存在的一种形式,我们可以认为某些病毒有化学结晶型和生命活动型的两种形式。
3.颗粒形式与基因形式病毒以颗粒形式存在于细胞之外,此时,只具感染性。一旦感染细胞病毒解体而释放出核酸基因组,然后才能进行复制和增殖,并产生新的子代病毒。有的病毒基因组整合于细胞基因组,随细胞的繁殖而增殖,此时病毒即以基因形式增殖,而不是以颗粒形式增殖,这是病毒潜伏感染的一种方式。
二、病毒结构和功能的两重性
1.标准病毒与缺陷病毒在病毒的增殖过程中,由于其基因组因某种微环境因素的影响或转录过程的错误而发生突变,以致有装配不全的病毒颗粒产生,称为缺陷病毒,产生缺陷病毒的原亲代病毒,则称为标准病毒,缺陷病毒颗粒有干扰标准病毒繁殖的作用。
2.假病毒与真病毒一种细胞有两种病毒同时感染的情况,在增殖过程中,一种病毒可以穿上本身的外壳,这就是真病毒,是这种病毒的应有“面目”;如果一种病毒的核酸被以另一病毒外壳来编码,则称为假病毒,此时一种病毒的本来性质,被另一种病毒的性质所掩盖。
3.杂种病毒和纯种病毒两种病毒混合感染时,除了出现假型病毒外,还有可能出现病毒核酸重组的情况,即一种病毒颗粒之中,可含有两种病毒的遗传物质,此可称为杂种病毒,这是病毒学中一个相当常见的现象。
三、病毒病理学的两重性
1.病毒的致病性和非致病性关于致病性和非致病性问题,是同宿主细胞相对而言的,在分子水平、细胞水平和机体水平,可能有不同的含义。在细胞水平有细胞病变作用,但在机体水平可能并不显示临床症状,此可称为亚临床感染或不显感染。
2.病毒感染的急性和慢性病毒感染所致的临床症状有急、慢之分,有的病毒一般只表现急性感染而很少表现慢性感染;有的则既有急性过程,也有慢性过程。
目前对病毒的概念可以是:病毒是代谢上无活性,有感染性,而不一定有致病性的因子,他们小于细胞,但大于大多数大分子,他们无例外地在生活细胞内繁殖,他们含有一个蛋白质或脂蛋白外壳和一种核酸,DNA或RNA,甚至只含有核酸而没有蛋白质,或只有蛋白质而没有核酸,它们作为大分子似乎太复杂,作为生物体它们的生理和复制方式又千姿百态。Lwoff在“病毒的概念”一文中强调病毒的特殊性时指出,“病毒应该就是病毒,因为它们是病毒”。
病毒_病毒 -历史发现
最早认知
人们最早是通过病毒导致的疾病认识到病毒的存在的。早在公元前二至三个世纪的印度和中国就有了关于天花的记录。但直到19世纪末,病毒才开始逐渐得以发现和鉴定。
最早记载的植物病毒病是郁金香是郁金香碎色病。
最早有记载的家畜中的病毒病是狂犬病。巴斯德(Pasteur)作为微生物发展史上的里程碑式的人物,因在1884年发明了狂犬疫苗,对病毒病的防治做出了巨大贡献。
科学研究
伊凡诺夫斯基发现植物病毒
1884年,法国微生物学家查理斯・尚柏朗(Charles Chamberland)发明了一种细菌无法滤过的过滤器(Chamberland氏烛形滤器,其滤孔孔径小于细菌的大小),他利用这一过滤器就可以将液体中存在的细菌除去。
1892年,俄国生物学家伊凡诺夫斯基(Dmitry Ivanovsky)在研究烟草花叶病时发现,将感染了花叶病的烟草叶的提取液用烛形滤器过滤后,依然能够感染其他烟草。于是他提出这种感染性物质可能是细菌所分泌的一种毒素,但他并未深入研究下去。当时,人们认为所有的感染性物质都能够被过滤除去并且能够在培养基中生长,这也是疾病的细菌理论(germ theory)的一部分。
1899年,荷兰微生物学家马丁乌斯・贝杰林克(Martinus Beijerinck)重复了Ivanovsky的实验,并相信这是一种新的感染性物质。他还观察到这种病原只在分裂细胞中复制,由于他的实验没有显示这种病原的颗粒形态,因此他称之为contagium vivum fluidum(可溶的活菌)并进一步命名为virus(病毒)。贝杰林克认为病毒是以液态形式存在的(但这一看法后来被温德尔・梅雷迪思・斯坦利推翻,他证明了病毒是颗粒状的)。同样在1899年,Friedrich Loeffler和Paul Frosch发现患口蹄疫动物淋巴液中含有能通过滤器的感染性物质,由于经过了高度的稀释,排除了其为毒素的可能性;他们推论这种感染性物质能够自我复制。
在19世纪末,病毒的特性被认为是感染性、可滤过性和需要活的宿主,也就意味着病毒只能在动物或植物体内生长。1906年,哈里森发明了在淋巴液中进行组织生长的方法;接着在1913年,E. Steinhardt、C. Israeli和R. A. Lambert利用这一方法在豚鼠角膜组织中成功培养了牛痘苗病毒,突破了病毒需要体内生长的限制。1928年,H. B. Maitland和M. C. Maitland有了更进一步的突破,他们利用切碎的母鸡肾脏的悬液对牛痘苗病毒进行了培养。他们的方法在1950年代得以广泛应用于脊髓灰质炎病毒疫苗的大规模生产。
另一项研究突破发生在1931年,美国病理学家Ernest William Goodpasture在受精的鸡蛋中培养了流感病毒。1949年,约翰・富兰克林・恩德斯、托马斯・哈克尔・韦勒和弗雷德里克・查普曼・罗宾斯利用人的胚胎细胞对脊髓灰质炎病毒进行了培养,这是首次在没有固体动物组织或卵的情况下对细菌进行的成功培养。这一研究成果被约纳斯・沙克利用来有效地生产脊髓灰质炎病毒疫苗。
1931年,德国工程师恩斯特・鲁斯卡和马克斯・克诺尔发明了电子显微镜,使得研究者首次得到了病毒形态的照片。1935年,美国生物化学家和病毒学家温德尔・梅雷迪思・斯坦利发现烟草花叶病毒大部分是由蛋白质所组成的,并得到病毒晶体。随后,他将病毒成功地分离为蛋白质部分和RNA部分。温德尔・斯坦利也因为他的这些发现而获得了1946年的诺贝尔化学奖。烟草花叶病毒是第一个被结晶的病毒,从而可以通过X射线晶体学的方法来得到其结构细节。第一张病毒的X射线衍射照片是由Bernal和Fankuchen于1941年所拍摄的。1955年,通过分析病毒的衍射照片,罗莎琳・富兰克林揭示了病毒的整体结构。同年,Heinz Fraenkel-Conrat和Robley Williams发现将分离纯化的烟草花叶病毒RNA和衣壳蛋白混合在一起后,可以重新组装成具有感染性的病毒,这也揭示了这一简单的机制很可能就是病毒在它们的宿主细胞内的组装过程。
20世纪早期,英国细菌学家Frederick Twort发现了可以感染细菌的病毒,并称之为噬菌体。随后法裔加拿大微生物学家Félix d'Herelle描述了噬菌体的特性:将其加入长满细菌的琼脂固体培养基上,一段时间后会出现由于细菌死亡而留下的空斑。高浓度的病毒悬液会使培养基上的细菌全部死亡,但通过精确的稀释,可以产生可辨认的空斑。通过计算空斑的数量,再乘以稀释倍数就可以得出溶液中病毒的个数。他们的工作揭开了现代病毒学研究的序幕。
20世纪的下半叶是发现病毒的黄金时代,大多数能够感染动物、植物或细菌的病毒在这数十年间被发现。1957年,马动脉炎病毒和导致牛病毒性腹泻的病毒(一种瘟病毒)被发现;1963年,巴鲁克・塞缪尔・布隆伯格发现了乙型肝炎病毒;1965年,霍华德・马丁・特明发现并描述了第一种逆转录病毒;这类病毒将RNA逆转录为DNA的关键酶,逆转录酶在1970年由霍华德・特明和戴维・巴尔的摩分别独立鉴定出来。1983年,法国巴斯德研究院的吕克・蒙塔尼和他的同事弗朗索瓦丝・巴尔-西诺西首次分离得到了一种逆转录病毒,也就是现在世人皆知的艾滋病毒(HIV)。其二人也因此与发现了能够导致子宫颈癌的人乳头状瘤病毒的德国科学家哈拉尔德・楚尔・豪森分享了2008年的诺贝尔生理学与医学奖。
病毒_病毒 -结构形状
病毒的形态各异,从简单的螺旋形和正二十面体形到复合型结构。人们在电镜下观察到许多病毒粒体的形态和大小,病毒的形状同其壳体的基本结构有着紧密的联系。病毒的壳体有四种结构类型,与之相对应,病毒颗粒的形状大致可分为下列四种类型:
螺旋对称壳体。蛋白质亚基沿中心轴呈螺旋状排列,形成高度有序、对称的稳定结构。螺旋对称的壳体形成直杆状、弯曲杆状和线状等杆状病毒颗粒。很多植物病毒如TMV等则为坚硬的杆状,而某些植物病毒和细菌病毒的形状是软而能弯曲的很长的纤维状。昆虫病毒中核型多角体病毒属成员也多呈杆状。
烟草花叶病毒结构:病毒RNA卷曲在的由重复的蛋白质亚基组成的螺旋形衣壳中。
二十面对称壳体。蛋白质亚基围绕具立方对称的正多面体的角或边排列,进而形成一个封闭的蛋白质的鞘。因二十面体容积为最大,能包装更多的病毒核酸,所以病毒壳体多取二十面体对称结构。病毒的壳体为二十面体对称,大部分动物病毒和少数植物病毒呈球状病毒颗粒。
具有正二十面体结构的腺病毒的电镜照片
复合对称壳体。仅少数病毒壳体为复合对称结构。壳体由头部和尾部组成,包装有病毒核酸的头部通常呈二十面体对称,尾部呈螺旋对称。具有复合对称结构的典型例子是有尾噬菌体(tailed phage)。有复合对称壳体的细菌病毒(噬菌体)和某些动物病毒呈复杂形状的病毒颗粒。
复合对称壳体
包膜型:一些病毒可以利用改造后的宿主的细胞膜(来自细胞表面的质膜或细胞内部的膜,如核膜及内质网膜)环绕在病毒体周围,形成一层脂质的包膜。包膜上既镶嵌有来自宿主的膜蛋白也有来自病毒基因组编码的膜蛋白;而脂质膜本身和其中的糖类则都来自宿主细胞。包膜型病毒位于包膜内的病毒体可以是螺旋形或正二十面体形的。
无包膜的病毒在宿主细胞内完成复制后,需要宿主细胞死亡并裂解后,才能逸出并进一步感染其他细胞。这种方法虽然简单,但常常造成大量非成熟细胞死亡,反而降低了对宿主细胞的利用率。而有了包膜之后,病毒可以通过包膜与宿主的细胞膜融合来出入细胞,而不需要造成细胞死亡。流感病毒和艾滋病毒就采用的是这种策略。大多数的包膜型病毒的感染性都依赖于包膜。
带状疱疹病毒的电镜负染照片显示其病毒颗粒周围被包膜所环绕
病毒_病毒 -化学组成
病毒
病毒的基本化学组成是核酸和蛋白质。所有成熟的病毒至少是由一种或几种蛋白质和一种核酸组成,只有少数几种例外,它们仅仅以核酸形式存在,如类病毒。有些病毒还含有一定量的脂类物质及碳水化合物,等等。一个病毒粒子是由DNA或RNA病毒核酸构成髓核。髓核被称之为衣壳的蛋白质外壳所包围。髓核和衣壳统称为核衣壳。有些病毒的核衣壳外面包被着一层囊膜,囊膜由脂质、蛋白质和糖组成。
病毒的核酸
核酸是病毒的遗传物质,携带着病毒的全部遗传信息,是病毒遗传和感染的物质基础。一种病毒的病毒颗粒只含有一种核酸,DNA或者RNA。它们以单链、双链或环状多核苷酸组成。一种病毒只有一种特定类型的核酸,DNA或RNA,这与某种特定类型的病毒起源有关。不同种类的病毒其核酸含量有较大的差别。流感病毒的核酸不到病毒颗粒质量的1%,大肠杆菌噬菌体T2、T4、T6的核酸约占病毒颗粒的一半或更多。由于核酸是病毒的遗传物质,每种病毒颗粒中的核酸含量并不一致,其结构和功能也有一定的关系。结构复杂的病毒有较多的核酸,结构简单的病毒只需较少的核酸。
病毒的蛋白质
蛋白质是病毒的另一类主要成分,包括结构蛋白和非结构蛋白。
非结构蛋白是指由病毒基因组编码的,在病毒复制或基因表达调控过程中具有一定功能,但不结合于病毒颗粒中的蛋白质。
结构蛋白是指构成一个形态成熟的有感染性的病毒颗粒所必需的蛋白质,包括壳体蛋白、包膜蛋白和毒粒酶等。
壳体蛋白是构成病毒壳体结构的蛋白质,由一条或多条多肽链折叠形成的蛋白质亚基,是构成壳体蛋白的最小单位。壳体蛋白的主要功能是:1)构成病毒的壳体,保护病毒的核酸。2)无包膜病毒的壳体蛋白参与病毒的吸附、侵入,决定病毒的宿主嗜性,同时它们还是病毒的表面抗原。
包膜蛋白是构成病毒包膜结构的蛋白质,包括包膜糖蛋白和基质蛋白两类。主要功能是:1)是病毒的主要表面抗原,它们与细胞受体相互作用启动病毒感染发生,有些还介导病毒的侵入。2)还可能具有凝集脊椎动物红血球细胞、细胞融合以及酶等活性。3)基质蛋白构成膜脂双层与核衣壳之间的亚膜结构,具有支撑包膜、维持病毒结构的作用,并在病毒芽出成熟过程中发挥重要作用。
毒粒酶根据功能大致分为两类:一类参与病毒侵入、释放等过程,如T4噬菌体的溶菌酶;一类参与病毒的大分子合成,如逆转录病毒的逆转录酶。
病毒的脂质
许多病毒的包膜内存在有脂类化合物,如磷脂、脂肪酸、甘油三酸脂和胆固醇等。这些脂类几乎都是由病毒粒子在细胞内成熟,在细胞膜处以葡生方式释放,直接从寄主细胞膜上得到。病毒脂类存在与病毒的吸附和侵入有关。
病毒的碳水化合物
除病毒的核酸中所含戊糖外,有的病毒还含有少量的碳水化合物,为核糖或脱氧核糖和磷酸组成的核酸骨架。有包膜病毒中碳水化合物以寡糖侧链的形式与蛋白质结合形成包膜糖蛋白。
其它组成
在某些动物、植物病毒中存在多胺类有机阳离子,包括丁二胺、亚精胺、精胺等,它们大都结合于病毒核酸,对核酸的构型有一定影响。在某些病毒的病毒体中,还发现有其它的小分子量组分,如ATP,对噬菌体尾鞘收缩提供能量。
病毒_病毒 -两重性
病毒生命形式的两重性
HIV病毒
1、病毒存在的两重性:病毒的生命活动很特殊,对细胞有绝对的依存性。其存在形式有二:一是细胞外形式,一是细胞内形式。存在于细胞外环境时,则不显复制活性,但保持感染活性,是病毒体或病毒颗粒形式。进入细胞内则解体释放出核酸分子(DNA或RNA),借细胞内环境的条件以独特的生命活动体系进行复制,是为核酸分子形式。
2、病毒的结晶性与非结晶性病毒可提纯为结晶体。某些病毒有化学结晶型和生命活动型的两种形式。
3、颗粒形式与基因形式:病毒以颗粒形式存在于细胞之外,此时,只具感染性。一旦感染细胞病毒解体而释放出核酸基因组,然后才能进行复制和增殖,并产生新的子代病毒。有的病毒基因组整合于细胞基因组,随细胞的繁殖而增殖,此时病毒即以基因形式增殖,而不是以颗粒形式增殖,这是病毒潜伏感染的一种方式。
病毒结构和功能的两重性
1、标准病毒与缺陷病毒:在病毒的增殖过程中,由于其基因组因某种微环境因素的影响或转录过程的错误而发生突变,以致有装配不全的病毒颗粒产生,称为缺陷病毒,产生缺陷病毒的原亲代病毒,则称为标准病毒,缺陷病毒颗粒有干扰标准病繁殖的作用。
2、假病毒与真病毒:一种细胞有两种病毒同时感染的情况,在增殖过程中,一种病毒可以穿上本身的外壳,这就是真病毒,是这种病毒的应有“面目”;如果一种病毒的核酸被以另一病毒编码的外壳,则称为假病毒,此时一种病毒的本来性质,被另一种病毒的性质所掩盖。
3、杂种病毒和纯种病毒:两种病毒混合感染时,除了出现假型病毒外,还有可能出现病毒核酸重组的情况,即一种病毒颗粒之中,可含有两种病毒的遗传物质,此可称为杂种病毒,这是病毒学中一个相当常见的现象。
病毒病理学的两重性
1、病毒的致病性和非致病性:关于致病性和非致病性问题,是同宿主细胞相对而言的,在分子水平、细胞水平和机体水平,可能有不同的含义。在细胞水平有细胞病变作用,但在机体水平可能并不显示临床症状,此可称为亚临床感染或不显感染。
2、病毒感染的急性和慢性:病毒感染所致的临床症状有急、慢之分,有的病毒一般只表现急性感染而很少表现慢性感染;有的则既有急性过程,也有慢性过程。
病毒_病毒 -生命周期
病毒
病毒缺乏增殖所需要的酶系统,只能在活的宿主细胞内增殖(自我复制)。绝大多数病毒复制过程可分为下列六步:吸附、侵入、脱壳、生物合成、组装和释放。
吸附
吸附是决定感染成功与否的关键环节。病毒吸附于敏感细胞需要病毒表面特异性的吸附蛋白与细胞表面受体相互作用。病毒吸附蛋白(virus attachment protein, VAP)一般由衣壳蛋白或包膜上的糖蛋白突起充当。细胞表面受体(也称为病毒受体,virus receptor)则为有效结合病毒粒子的细胞表面结构,大多数噬菌体的病毒受体为细菌细胞壁上的磷壁酸分子、脂多糖分子以及糖蛋白复合物,有的则位于菌毛、鞭毛或荚膜上。大部分动物病毒的病毒受体为镶嵌在细胞膜脂质双分子层中的糖蛋白,也有的是糖脂或唾液酸寡糖苷。植物病毒迄今尚未发现有特异性细胞受体,其进入植物细胞的机制是通过伤口或媒介传播。
病毒的细胞受体具有种系和组织特异性,决定了病毒的宿主谱。不同种属的病毒其细胞受体不同,有的甚至同种不同型的病毒以及同型不同株的病毒受体也不相同;另一方面,有些不同种属的病毒却有相同的细胞受体,其吸附和感染可对其它病毒的感染产生干扰。
VAP与病毒受体的结合需要一定的温度条件,以促进与酶反应相类似的化学反应。在0-37℃内温度越高病毒吸附效率也越高。病毒吸附细胞的过程可在几分钟到几十分钟的时间内完成。
侵入
病毒通过以下不同的方式进入宿主细胞:注射式侵入、细胞内吞、膜融合以及其它特殊的侵入方式。
注射式侵入是有尾噬菌体通常的侵入方式。通过尾部收缩将衣壳内的DNA基因组注入宿主细胞内。
细胞内吞是动物病毒的常见侵入方式。经细胞膜内陷形成吞噬泡,使病毒粒子进入细胞质中。
膜融合是有包膜病毒侵入过程中,病毒包膜与细胞膜融合的一种侵入方式。
直接侵入大致可分为几种类型。1)部分病毒粒子直接侵入宿主细胞,其机理不明;2)病毒与细胞膜表面受体结合后,由细胞表面的酶类帮助病毒粒体释放核酸进入细胞质中,病毒衣壳仍然留在细胞膜外,将病毒侵入和脱壳融为一体。3)其它特殊方式。植物病毒通过存在于植物细胞壁上的小伤口或天然的外壁孔侵入,或植物细胞之间的胞间连丝侵入细胞,也可通过介体的口器、吸器等侵入细胞。
脱壳
脱壳是指病毒感染性核酸从衣壳内释放出来的过程。有包膜病毒脱壳包括脱包膜和脱衣壳两个步骤,无包膜病毒只需脱衣壳,方式随不同病毒而异。
注射式侵入的噬菌体和某些直接侵入的病毒可以直接在细胞膜或细胞壁表面同步完成侵入和脱壳。病毒粒子以内吞方式或直接进入细胞后,经蛋白酶的降解,先后脱去包膜和衣壳。以膜融合方式侵入的病毒,其包膜在与细胞膜融合时即已脱掉,核衣壳被移至脱壳部位并在酶的作用下进一步脱壳,病毒核酸游离并进至细胞的一定部位进行生物合成。病毒脱壳必须有酶的参与,脱壳酶来自宿主细胞,有的为病毒基因编码。
生物合成
病毒借助宿主细胞提供的原料、能量和场所合成核酸和蛋白质,期间所需的多数酶也来自宿主细胞。在病毒进入宿主细胞后生物合成阶段,胞浆中无病毒颗粒,称为隐蔽期(eclipse)。
装配
病毒的结构成分核酸与蛋白质分别合成后,在细胞核内或细胞质内组装成核衣壳。绝大多数DNA病毒在细胞核内组装,RNA病毒与痘病毒类则在细胞质内组装。无包膜病毒组装成核衣壳即为成熟的病毒体,病毒的早期蛋白,即非病毒结构成分不组装入病毒,残留在感染细胞中。
释放
绝大多数无包膜病毒释放时被感染的细胞崩解,释放出病毒颗粒,宿主细胞膜破坏,细胞迅即死亡。绝大多数有包膜病毒通过细胞内的内质网、空泡,或包上细胞核膜或细胞膜以出芽方式释放而成为成熟病毒,在一段时间内逐个释出,对细胞膜破坏轻,宿主细胞死亡慢。从单个病毒吸附开始至所有病毒释放,此过程称为感染周期或复制周期。一个感染细胞一般释放的病毒数为100-1000。
病毒_病毒 -遗传变异
遗传受阻
脊髓灰质炎病毒病毒对敏感细胞的感染并不一定都能繁殖病毒,产生有感染性的病毒子代。由于病毒或细胞的原因,使病毒的复制在病毒进入敏感细胞后的某一阶段受阻,导致病毒感染的不完全循环,不产生有感染性的病毒子代。
缺损病毒:有些病毒由于缺乏某些基因,单独感染细胞时不能复制出完整的、具有感染性的病毒颗粒,需要其它病毒基因组或病毒基因的辅助活性,否则,即使在活细胞内也不能复制。
顿挫病毒:有些宿主细胞不能全部提供病毒复制所需的必要因子,致使所复制的病毒为不完整的、无感染性的病毒颗粒或亚颗粒。
干扰现象:两种病毒感染同一种细胞或机体时,常常发生一种病毒抑制另一种病毒复制的现象,称为干扰现象(interference)。干扰现象可在同种以及同株的病毒间发生,后者如流感病毒的自身干扰。异种病毒和无亲缘关系的病毒之间也可以干扰,且比较常见。
变异遗传
病毒的遗传变异可以有多种机制。“遗传漂变”(geneticdrift)是其中之一,即病毒DNA或RNA上单个碱基的突变。大多数这样的单点突变是无义的(或者说是沉默的),因为它们没有导致所编码的蛋白质发生变化;但有一小部分突变可能会引起进化上的优势,如产生对抗病毒药物的抵抗力。抗原转移(antigenicshift)是另一种病毒基因组的主要变化,是由遗传重组或基因重排所导致的。当流感病毒发生抗原转移后,可能会导致瘟疫。RNA病毒常常以准种(quasispecies)的形式或大量同种但基因组核苷酸序列存在微小差异的病毒的形式存在。这样的准种是自然选择的主要目标。
遗传重组是一条DNA链断裂后重新连接到另一条不同DNA分子末端的过程。遗传重组可以在病毒感染细胞的同时发生。对病毒进化的研究结果显示,在已研究的各种病毒中,重组发生得极为频繁。而且,无论是RNA病毒还是DNA病毒,重组的发生都是非常普遍的。
病毒_病毒 -灭活
病毒受理化因素作用后失去感染性,称为灭活(inactivation)。灭活的病毒仍保留其抗原性、红细胞吸附、血凝和细胞融合等活性。
物理因素
1)温度。大多数病毒耐冷不耐热,在0℃以下温度能良好生存,特别是在干冰温度(-70℃)和液氮温度(-196℃)下更可长期保持其感染性;相反,大多数病毒于55-60℃下,几分钟至十几分钟即被灭活,100℃时在几秒钟内即可灭活病毒。
2)pH。一般来说,大多数病毒在pH 6-8的范围内比较稳定,而在pH5.0以下或者pH9.0以上容易灭活。
3)辐射。电离辐射中的Υ射线和X射线以及非电离辐射中的紫外线都能使病毒灭活。
化学因素
1)脂溶剂。有包膜病毒对脂溶剂敏感。乙醚、氯仿、丙酮、阴离子去垢剂等均可使有包膜病毒灭活。借此可以鉴别有包膜病毒和无包膜病毒。
2)氧化剂、卤素、醇类。病毒对各种氧化剂、卤素、醇类物质敏感。H2O2、漂白粉、高锰酸钾、甲醛、过氧乙酸、次氯酸盐、酒精、甲醇等均可灭活病毒。
3)抗生素和中草药。病毒对抗生素不敏感,在病毒分离时,标本用抗生素处理或在培养液中加入抗生素可抑制标本中的杂菌,有利于病毒分离。近年来的研究表明,有些中药如板蓝根、大青叶、柴胡、大黄、贯仲等对某些病毒有抑制作用。
病毒_病毒 -分类
由于病毒并不像其他生物能借由交配产生后代,因此在种别的定义上与一般生物有所不同。ICTV分类法
巴尔的摩分类法是基于病毒mRNA的合成方式
国际病毒分类委员会(International Committee on Taxonomy of Viruses, 简称ICTV)在1966年建立起了一个病毒分类的通用系统和统一的命名法则。其七届ICTV会议首次规范化了病毒物种的概念,即病毒分类的分支层次中的最低分类单元。分类的主要依据是病毒颗粒的特性、抗原特性与生物特性。
分类结构如下:
目 (-virales)
科 (-viridae)
亚科(-virinae)
属 (-virus)
种 (-virus)
在2008年的ICTV分类中,五个目已经建立,分别是有尾噬菌体目(Caudovirales)、Herpesvirales、单股反链病毒目(Mononegavirales)、Nidovirales和Picornavirales。分类委员会没有正式区分亚种、株系和分离株之间的区别。分类表中总共有5个目、82个科、11个亚科、307个属、2083个种以及约3000种尚未分类的病毒类型。
巴尔的摩分类法
诺贝尔奖获得者生物学家戴维・巴尔的摩在1970年代提出了巴尔的摩分类系统。巴尔的摩分类法是基于病毒mRNA的生成机制。在从病毒基因组到蛋白质的过程中,必须要生成mRNA来完成蛋白质合成和基因组的复制,但每一个病毒家族都采用不同的机制来完成这一过程。病毒基因组可以是单链或双链的RNA或DNA,可以有也可以没有反转录酶。而且,单链RNA病毒可以是正义( )或反义(-)。这一分类法将病毒分为7类:
第一类是双链DNA病毒(如腺病毒、疱疹病毒、痘病毒)
第二类是单链DNA病毒(如小DNA病毒)
第三类是双链RNA病毒(如呼肠孤病毒)
第四类是( )单链RNA病毒(如微小核糖核酸病毒、披盖病毒)
第五类是(?)单链RNA病毒(如正黏液病毒、炮弹病毒)
第六类是单链RNA反转录病毒(如反转录病毒)
第七类是双链DNA反转录病毒(如肝病毒)
举一个病毒分类的例子:水痘病毒,即带状疱疹病毒,属于Herpesvirales目,疱疹病毒科,甲型疱疹病毒亚科,水疱病毒属;同时,带状疱疹病毒是巴尔的摩分类法中的第一类,因为它是双链DNA病毒,且不含有反转录酶。
病毒由于不能独立生活,必须生活在其他生物的细胞内。根据他们寄生的细胞不同,可以将病毒分为三大类:专门寄生在人和动物细胞里面的动物病毒;专门寄生在植物细胞里的植物病毒,如烟草花叶病毒;专门寄生在细菌细胞内的细菌病毒,也叫噬菌体,如大肠杆菌噬菌体。
病毒_病毒 -传播方式
巴斯德(Pasteur)
病毒的传播方式多种多样,不同类型的病毒采用不同的方法。例如,植物病毒可以通过以植物汁液为生的昆虫,如蚜虫,来在植物间进行传播;而动物病毒可以通过蚊虫叮咬而得以传播。这些携带病毒的生物体被称为“载体”。流感病毒可以经由咳嗽和打喷嚏来传播;诺罗病毒则可以通过手足口途径来传播,即通过接触带有病毒的手、食物和水;轮状病毒常常是通过接触受感染的儿童而直接传播的;此外,艾滋病毒则可以通过性接触来传播。
并非所有的病毒都会导致疾病,因为许多病毒的复制并不会对受感染的器官产生明显的伤害。一些病毒,如艾滋病毒,可以与人体长时间共存,并且依然能保持感染性而不受到宿主免疫系统的影响,即“病毒持续感染”(viral persistence)。但在通常情况下,病毒感染能够引发免疫反应,消灭入侵的病毒。而这些免疫反应能够通过注射疫苗来产生,从而使接种疫苗的人或动物能够终生对相应的病毒免疫。像细菌这样的微生物也具有抵御病毒感染的机制,如限制修饰系统。抗生素对病毒没有任何作用,但抗病毒药物已经被研发出来用于治疗病毒感染。
病毒_病毒 -引起疾病
天花病毒1、与人类相关的疾病
由病毒引起的人类疾病种类繁多。已经确定的如,伤风、流感、水痘等一般性疾病,以及天花、艾滋病、SARS和禽流感等严重疾病。还有一些疾病可能是以病毒为致病因子;例如,人疱疹病毒6型与一些神经性疾病,如多发性硬化症和慢性疲劳综合症之间可能相关。此外,原本被认为是马的神经系统疾病的致病因子的玻那病毒,现在被发现可能能够引起人类精神疾病。病毒能够导致疾病的能力被称为病毒性(virulence)。
一些病毒能够引起慢性感染,可以在机体内不断复制而不受宿主防御系统的影响。这类病毒包括乙肝病毒和丙肝病毒。
2、与其他生物体相关的疾病禽流感病毒
病毒可以感染所有的物种,少数病毒(如mimivirus)甚至也会受到其他特定病毒的感染;但特定的病毒感染物种的范围是有限的。例如,植物病毒不会感染动物,而噬菌体只能感染细菌。
1、动物
对家畜来说,病毒是重要的致病因子;能够导致的疾病包括口蹄疫、蓝舌病等。作为人类宠物的猫、狗、马等,如果没有接种疫苗,会感染一些致命病毒。例如犬小病毒(Canine parvovirus),一种小DNA病毒,其感染是导致幼犬死亡的重要原因。所有的无脊椎动物都会感染病毒。
2、植物
植物病毒的种类繁多,能够影响受感染植物的生长和繁殖。植物病毒的传播常常是由被称为“载体”的生物来完成。这些载体一般为昆虫,也有部分情况下为真菌、线虫动物以及一些单细胞生物。控制针对植物的病毒感染,通常是采用消灭载体生物以及除去其他可能的病毒宿主,如杂草。对于人类及其他动物来说,植物病毒是无害的,因为它们只能够在活的植物细胞内进行复制。植物具备精巧而有效的防御机制来抵抗病毒感染。
3、细菌
噬菌体是病毒中最为普遍和分布最广的群体。
4、古菌
古菌也会被一些病毒感染,主要是双链DNA病毒。
病毒_病毒 -应用
棉铃虫核型多角体病毒杀虫剂一、生命科学与医学
病毒对于分子生物学和细胞生物学的研究具有重要意义,因为它们提供了能够被用于改造和研究细胞功能的简单系统。研究和利用病毒为细胞生物学的各方面研究提供了大量有价值的信息。例如,病毒被用在遗传学研究中来帮助我们了解分子遗传学的基本机制,包括DNA复制、转录、RNA加工、翻译、蛋白质转运以及免疫学等。
二、材料科学与纳米技术纳米技术的发展趋势是制造多用途的病毒。从材料科学的观点来看,病毒可以被看作有机纳米颗粒:它们的表面携带特定的工具用于穿过宿主细胞的壁垒。病毒的大小和形状,以及它们表面的功能基团的数量和性质,是经过精确地定义的。正因为如此,病毒在材料科学中被普遍用作支架来共价连接表面修饰。病毒的一个特点是它们能够通过直接进化来被改动。从生命科学发展而来的这些强大技术正在成为纳米材料制造方法的基础,远远超越了它们在生物学和医学中的应用而被应用于更加广泛的领域中。
三、武器病毒能够引起瘟疫而导致人类社会的恐慌,这种能力使得一些人企图利用病毒作为生化武器来达到常规武器所不能获得的效果。而随着臭名昭著的西班牙流感病毒在实验室中获得成功复原,对于病毒成为武器的担心不断增加。
四、其他用途1、噬菌体可以作为防治某些疾病的特效药,例如烧伤病人在患处涂抹绿浓杆菌噬菌体稀释液。
2、在细胞工程中,某些病毒可以作为细胞融合的助融剂,例如仙台病毒。
3、在基因工程中,病毒可以作为目的基因的载体,使之被拼接在目标细胞的染色体上。
4、在专一的细菌培养基中添加的病毒可以除杂。
5、病毒可以作为精确制导药物的载体。
6、病毒可以作为特效杀虫剂。
7、病毒还在生物圈的物质循环和能量交流中起到关键作用。
8、利用灭活制作病毒疫苗。
病毒_病毒 -起源说
艾滋病病毒
有三种流行的关于病毒起源的理论:
1、逆向理论(Regressive theory):病毒可能曾经是一些寄生在较大细胞内的小细胞。随着时间的推移,那些在寄生生活中非必需的基因逐渐丢失。这一理论的证据是,细菌中的立克次氏体和衣原体就像病毒一样,需要在宿主细胞内才能复制;而它们缺少了能够独立生活的基因,这很可能是由于寄生生活所导致的。这一理论又被称为退化理论(degeneracy theory)。2、细胞起源理论(有时也称为漂荡理论):一些病毒可能是从较大生物体的基因中“逃离”出来的DNA或RNA进化而来的。逃离的DNA可能来自质粒(可以在细胞间传递的裸露DNA分子)或转座子(可以在细胞基因内不同位置复制和移动的DNA片断,曾被称为“跳跃基因”,属于可移动遗传元件)。转座子是在1950年由巴巴拉・麦克林托克在玉米中发现的。
3、共进化理论:病毒可能进化自蛋白质和核酸复合物,与细胞同时出现在远古地球,并且一直依赖细胞生命生存至今。类病毒是一类RNA分子,但不被归入病毒中,因为它们缺少由蛋白质形成的衣壳。然而,它们具有多种病毒的普遍特征,常常被称为亚病毒物质。类病毒是重要的植物病原体。它们没有编码蛋白质的基因,但可以与宿主细胞作用,利用宿主来进行它们自身的复制。这些依赖于其他种类病毒的病毒被称为“卫星病毒”,它们可能是介于类病毒和病毒之间的进化中间体。
病毒_病毒 -第一个发现癌的人
弗朗西斯・佩顿・劳斯(FrancisPeytonRous)1879年10月5日出生于美国,是纽约市洛克菲勒研究所的内科医生和病毒学家。 劳斯医生毕业于马里兰州巴尔的摩市约翰斯・霍普金斯大学。1911年1月21日,弗朗西斯・佩顿・劳斯发表了一份报告:癌性肿瘤是病毒所致。这一提法在医学史上是首次。因为还没有证据表明癌症对人或动物有传染性。劳斯也成为发现这种“肿瘤病毒”的第一人,因为这种病毒最先是在那只被劳斯接诊的鸡身上发现的,所以病毒被命名为“劳斯鸡肉瘤病毒”。1966年,已经87岁高龄的劳斯在距离发现这种病毒55年之后,获得了诺贝尔生理学或医学奖。这种病毒的发现与劳斯积极的工作是分不开的,多年来,劳斯一直在积极地进行着研究工作,事实上,直到他过90岁生日时为止,他一直都在工作。
病毒_病毒 -功能性治愈艾滋婴儿病毒
美国研究人员3日报告说,他们通过抗逆转录病毒疗法实现了首次“功能性治愈”艾滋病病毒婴儿感染者。
美国约翰斯・霍普金斯儿童医疗中心、密西西比大学等机构的研究人员3日在美国亚特兰大举行的“2013年逆转录病毒与机会性感染大会”上报告说,他们在两年前选取一名通过母婴传播感染艾滋病病毒的女婴为治疗对象。在该婴儿出生30小时后,研究人员对其进行组合式抗逆转录病毒治疗。检测证明,经过治疗该女婴血液中艾滋病病毒的数量明显递减,在其出生29天后,体内的艾滋病病毒已经检测不到。研究人员在随后18个月内继续对她进行抗逆转录治疗,并在停止治疗10个月后发现,其体内的艾滋病病毒抗体仍为阴性,常规血液检测中未发现艾滋病病毒存在。
研究人员说:“对新生儿进行抗逆转录病毒治疗后可以阻止体内藏匿的艾滋病病毒感染宿主细胞,该疗法能够清除、抑制该病毒,在非终身治疗的情况下实现‘功能性治愈’。”
“功能性治愈”是指感染者体内的艾滋病病毒被完全抑制,机体免疫功能正常,即便不接受治疗,用常规方法也难以在患儿血液中检测出病毒。 研究人员表示,根除艾滋病病毒,即“根本性治愈”艾滋病当前难以实现。针对艾滋病病毒婴儿感染者的药物治疗,一般开始于其出生后3到4个月之间,因此尽早、准确进行抗逆转录治疗对感染艾滋病病毒的婴儿意义重大。